Placenta As a Source of Hematopoietic Stem Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vocabulario De Morfoloxía, Anatomía E Citoloxía Veterinaria
Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) Servizo de Normalización Lingüística Universidade de Santiago de Compostela COLECCIÓN VOCABULARIOS TEMÁTICOS N.º 4 SERVIZO DE NORMALIZACIÓN LINGÜÍSTICA Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) 2008 UNIVERSIDADE DE SANTIAGO DE COMPOSTELA VOCABULARIO de morfoloxía, anatomía e citoloxía veterinaria : (galego-español- inglés) / coordinador Xusto A. Rodríguez Río, Servizo de Normalización Lingüística ; autores Matilde Lombardero Fernández ... [et al.]. – Santiago de Compostela : Universidade de Santiago de Compostela, Servizo de Publicacións e Intercambio Científico, 2008. – 369 p. ; 21 cm. – (Vocabularios temáticos ; 4). - D.L. C 2458-2008. – ISBN 978-84-9887-018-3 1.Medicina �������������������������������������������������������������������������veterinaria-Diccionarios�������������������������������������������������. 2.Galego (Lingua)-Glosarios, vocabularios, etc. políglotas. I.Lombardero Fernández, Matilde. II.Rodríguez Rio, Xusto A. coord. III. Universidade de Santiago de Compostela. Servizo de Normalización Lingüística, coord. IV.Universidade de Santiago de Compostela. Servizo de Publicacións e Intercambio Científico, ed. V.Serie. 591.4(038)=699=60=20 Coordinador Xusto A. Rodríguez Río (Área de Terminoloxía. Servizo de Normalización Lingüística. Universidade de Santiago de Compostela) Autoras/res Matilde Lombardero Fernández (doutora en Veterinaria e profesora do Departamento de Anatomía e Produción Animal. -

Messages from the Placentae Across Multiple Species a 50 Years
Placenta 84 (2019) 14–27 Contents lists available at ScienceDirect Placenta journal homepage: www.elsevier.com/locate/placenta Messages from the placentae across multiple species: A 50 years exploration T Hiroaki Soma Saitama Medical University, Japan ARTICLE INFO ABSTRACT Keywords: This review explores eight aspects of placentation in multiple mammalian. Gestational trophoblastic disease 1) Specialities of gestational trophoblastic disease. SUA(Single umbilical artery) 2) Clinical significance of single umbilical artery (SUA) syndrome. DIC(Disseminated intravascular coagulation) in 3) Pulmonary trophoblast embolism in pregnant chinchillas and DIC in pregnant giant panda. giant panda 4) Genetics status and placental behaviors during Japanese serow and related antelopes. Placentation in Japanese serow 5) Specific living style and placentation of the Sloth and Proboscis monkey. Hydatidiform mole in chimpanzee Placentation in different living elephant 6) Similarities of placental structures between human and great apes. Manatee and hyrax 7) Similarities of placental forms in elephants, manatees and rock hyrax with different living styles. Specific placental findings of Himalayan people 8) Specialities of placental pathology in Himalayan mountain people. Conclusions: It was taught that every mammalian species held on placental forms applied to different environ- mental life for their infants, even though their gestational lengths were different. 1. Introduction of effective chemotherapeutic agents. In 1959, I was fortunate tore- ceive an invitation from Prof. Kurt Benirschke at the Boston Lying-in Last October, Scientific American published a special issue about a Hospital. Before that, I had written to Prof. Arthur T. Hertig, Chairman baby's first organ, the placenta [1]. It is full of surprises and amazing of Pathology, Harvard Medical School, asking to study human tropho- science. -

FASD Effects of Alcohol on a Fetus
EFFECTS OF ALCOHOL ON A FETUS “Of all the substances of abuse (including cocaine, heroin, and marijuana), alcohol produces by far the most serious neurobehavioral effects in the fetus.” —Institute of Medicine Report to Congress, 19961 Prenatal exposure to alcohol can damage a fetus at any time, causing problems that persist throughout the individual’s life. There is no known safe level of alcohol use in pregnancy. WHAT IS THE SCOPE OF THE PROBLEM? identified in virtually every part of the body, including the brain, face, eyes, ears, heart, kidneys, and bones. No Alcohol is one of the most dangerous teratogens, which single mechanism can account for all the problems that are substances that can damage a developing fetus.1 Every alcohol causes. Rather, alcohol sets in motion many time a pregnant woman has a drink, her unborn child processes at different sites in the developing fetus: has one, too. Alcohol, like carbon monoxide from cigarettes, passes easily through the placenta from the • Alcohol can trigger cell death in a number of ways, mother's bloodstream into her baby's blood (See Figure causing different parts of the fetus to develop abnormally. 1)—and puts her fetus at risk of having a fetal alcohol • Alcohol can disrupt the way nerve cells develop, travel spectrum disorder (FASD). The blood alcohol level to form different parts of the brain, and function. (BAC) of the fetus becomes equal to or greater than the blood alcohol level of the mother. Because the fetus • By constricting the blood vessels, alcohol interferes with cannot break down alcohol the way an adult can, its BAC blood flow in the placenta, which hinders the delivery 2 remains high for a longer period of time. -
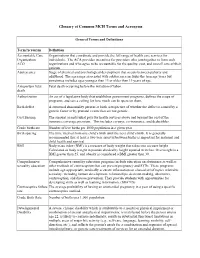
Glossary of Common MCH Terms and Acronyms
Glossary of Common MCH Terms and Acronyms General Terms and Definitions Term/Acronym Definition Accountable Care Organizations that coordinate and provide the full range of health care services for Organization individuals. The ACA provides incentives for providers who join together to form such ACO organizations and who agree to be accountable for the quality, cost, and overall care of their patients. Adolescence Stage of physical and psychological development that occurs between puberty and adulthood. The age range associated with adolescence includes the teen age years but sometimes includes ages younger than 13 or older than 19 years of age. Antepartum fetal Fetal death occurring before the initiation of labor. death Authorization An act of a legislative body that establishes government programs, defines the scope of programs, and sets a ceiling for how much can be spent on them. Birth defect A structural abnormality present at birth, irrespective of whether the defect is caused by a genetic factor or by prenatal events that are not genetic. Cost Sharing The amount an individual pays for health services above and beyond the cost of the insurance coverage premium. This includes co-pays, co-insurance, and deductibles. Crude birth rate Number of live births per 1000 population in a given year. Birth spacing The time interval from one child’s birth until the next child’s birth. It is generally recommended that at least a two-year interval between births is important for maternal and child health and survival. BMI Body mass index (BMI) is a measure of body weight that takes into account height. -

What Is the Role of the Placenta—Does It Protect Against Or Is It a Target for Insult?
Teratology Primer, 3rd Edition www.teratology.org/primer What Is the Role of the Placenta—Does It Protect Against or Is It a Target for Insult? Richard K. Miller University of Rochester School of Medicine & Dentistry Rochester, New York The placenta is not just a barrier but has many functions that are vital to the health of the embryo/fetus. The placenta is the anchor, the conduit, and the controller of pregnancy—and it can also be a target for toxicant action. The placenta encompasses not only the chorioallantoic placenta but all of its extraembryonic membranes (chorion/amnion) and the yolk sac. (Figure 1). The placenta and its membranes secure the embryo and fetus to the decidua (uterine lining) and release a variety of steroid and protein hormones that characterize the physiology of the pregnant female. Figure 1. Placental Structure in the Mouse. Figures depict early development of the mouse conceptus at embryonic days (E3.5, E7.5, E12.5). In the fetus, the visceral yolk sac (vYS) inverts and remains active throughout the entire gestation providing for transfer of selective large molecules, e.g., immunoglobulins IgG and vitamin B12. Abbreviations: Al, allantois; Am, amnion; Ch, chorion; Dec, decidua; Emb, embryo; Epc, ectoplacental cone; ICM, inner cell mass; Lab, Labyrinth; pYS, parietal yolk sac; SpT, spongiotrophoblast; TCG, trophoblast giant cell; Umb Cord, umbilical cord; vYS, visceral yolk sac; C-TGC, maternal blood canal trophoblast giant cell; P-TGC, parietal trophoblast giant cell; S-TGC, sinusoidal trophoblast giant cell; SpA-TGC, Spiral artery-associated trophoblast giant cell; Cyan-trophectoderm and trophoblast lineage, Black- inner cell mass and embryonic ectoderm; Gray -endoderm, Red-maternal vasculature, Purple-mesoderm, Yellow- decidua, Pink-fetal blood vessels in labyrinth. -

Equine Placenta – Marvelous Organ and a Lethal Weapon
Equine placenta – marvelous organ and a lethal weapon Malgorzata Pozor, DVM, PhD, Diplomate ACT Introduction Placenta has been defined as: „an apposition between parent (usually maternal) and fetal tissue in order to establish physiological exchange” (1). Another definition of this important organ was proposed by Steven and Morris: „a device consisting of one or more transport epithelia located between fetal and maternal blood supply” (2). The main function of placenta is to provide an interface between the dam and the the fetus and to allow the metabolic exchange of the the nutrients, oxygen and waste material. The maternal circulation is brought into a close apposition to the fetal circulation, while a separation of these two circulatory systems remain separated (3). A degree and complexity of this „intimate relationship” varies greately between species mostly due to the structural diversity of the extraembryonic membranes of the vertebrates. The early feto-maternal exchange in the equine pregnancy is established as early as on day 22 after fertilization. The fetal and choriovitellin circulations are already present, the capsule ruptures and the allantois is already visible (4). The allantois starts expanding by day 32 and vascularizes approximately 90% of the chorion and fuses with it to form chorioallantois by day 38 of gestation (5). The equine placenta continues increasing its complexity till approximately day 150 of gestation. Equids have epitheliochorial placenta, there are six leyers separating maternal and fetal circulation, and there are no erosion of the luminal, maternal epithelium, like in ruminants (6). Thousands of small chorionic microvilli develop and penetrate into endometrial invaginations. -

4 Extraembryonic Membranes
Implantation, Extraembryonic Membranes, Placental Structure and Classification A t t a c h m e n t and Implantation Implantation is the first stage in development of the placenta. In most cases, implantation is preceded by a close interaction of embryonic trophoblast and endometrial epithelial cells that is known as adhesion or attachment. Implantation also is known as the stage where the blastocyst embeds itself in the endometrium, the inner membrane of the uterus. This usually occurs near the top of the uterus and on the posterior wall. Among other things, attachment involves a tight intertwining of microvilli on the maternal and embryonic cells. Following attachment, the blastocyst is no longer easily flushed from the lumen of the uterus. In species that carry multiple offspring, attachment is preceeded by a remarkably even spacing of embryos through the uterus. This process appears to result from uterine contractions and in some cases involves migration of embryos from one uterine horn to another (transuterine migration). The effect of implantation in all cases is to obtain very close apposition between embryonic and maternal tissues. There are, however, substantial differences among species in the process of implantation, particularly with regard to "invasiveness," or how much the embryo erodes into maternal tissue. In species like horses and pigs, attachment and implantation are essentially equivalent. In contrast, implantation in humans involves the embryo eroding deeply into the substance of the uterus. •Centric: the embryo expands to a large size before implantation, then remains in the center of the uterus. Examples include carnivores, ruminants, horses, and pigs. •Eccentric: The blastocyst is small and implants within the endometrium on the side of the uterus, usually opposite to the mesometrium. -

From Trophoblast to Human Placenta
From Trophoblast to Human Placenta (from The Encyclopedia of Reproduction) Harvey J. Kliman, M.D., Ph.D. Yale University School of Medicine I. Introduction II. Formation of the placenta III. Structure and function of the placenta IV. Complications of pregnancy related to trophoblasts and the placenta Glossary amnion the inner layer of the external membranes in direct contact with the amnionic fluid. chorion the outer layer of the external membranes composed of trophoblasts and extracellular matrix in direct contact with the uterus. chorionic plate the connective tissue that separates the amnionic fluid from the maternal blood on the fetal surface of the placenta. chorionic villous the final ramification of the fetal circulation within the placenta. cytotrophoblast a mononuclear cell which is the precursor cell of all other trophoblasts. decidua the transformed endometrium of pregnancy intervillous space the space in between the chorionic villi where the maternal blood circulates within the placenta invasive trophoblast the population of trophoblasts that leave the placenta, infiltrates the endo– and myometrium and penetrates the maternal spiral arteries, transforming them into low capacitance blood channels. Sunday, October 29, 2006 Page 1 of 19 From Trophoblasts to Human Placenta Harvey Kliman junctional trophoblast the specialized trophoblast that keep the placenta and external membranes attached to the uterus. spiral arteries the maternal arteries that travel through the myo– and endometrium which deliver blood to the placenta. syncytiotrophoblast the multinucleated trophoblast that forms the outer layer of the chorionic villi responsible for nutrient exchange and hormone production. I. Introduction The precursor cells of the human placenta—the trophoblasts—first appear four days after fertilization as the outer layer of cells of the blastocyst. -

Terminologia Embryologica Y Placenta: Propuesta De Términos Embriológicos En Español
Int. J. Morphol., 36(1):63-68, 2018. Terminologia Embryologica y Placenta: Propuesta de Términos Embriológicos en Español Terminologia Embryologica and Placenta: Proposal of Embryological Terms in Spanish Ruth Prieto Gómez1 & Nicolás Ernesto Ottone2,3 PRIETO, G. R. & OTTONE, N. E. Terminologia Embryologica y placenta: Propuesta de Términos Embriológicos en español. Int. J. Morphol., 36(1):63-68, 2018. RESUMEN: En el área de la embriología, y en relación al uso de Terminologia Embryologica (TE), existen términos que son utilizados y que no se corresponden con ésta última. Pero a esta situación clásica, desde el origen de Nomina Anatomica de Basilea en 1895, se suma la ausencia de términos embriológicos en TE y que son diariamente reconocidos y nombrados en la práctica clínica. Además, no existe aún traducción oficial al español de TE. El objetivo de este trabajo consistió en realizar una propuesta de términos en español correspondientes a los términos incluídos en Paraplacenta [E6.0.2.4.0.1.], Placenta [E5.11.3.1.1.0.5] y Anomaliae placentae [E6.0.2.5.1.0.1], a partir de Terminologia Embryologica (TE) publicada por el Federal International Programme on Anatomical Terminologies en 2013, y en la cual sólo se encuentra la traducción al idioma inglés. La importancia de todos los trabajos relacionados con el buen uso de las terminologías y su correcta traducción al idioma vernáculo, radica en que la aplicación de un lenguaje único y común permitirá una mejor y mayor difusión de las investigaciones en el área de las ciencias morfológicas. PALABRAS CLAVE: Terminologia Embryologica; Placenta. -

The Allantois and Chorion, When Isolated Before Circulation Or Chorio-Allantoic Fusion, Have Hematopoietic Potential
Dartmouth College Dartmouth Digital Commons Open Dartmouth: Published works by Dartmouth faculty Faculty Work 11-2006 The Allantois and Chorion, when Isolated before Circulation or Chorio-Allantoic Fusion, have Hematopoietic Potential Brandon M. Zeigler Dartmouth College Daisuke Sugiyama Dartmouth College Michael Chen Dartmouth College Yalin Guo Dartmouth College K. M. Downs University of Wisconsin-Madison See next page for additional authors Follow this and additional works at: https://digitalcommons.dartmouth.edu/facoa Part of the Biochemistry Commons, Cell and Developmental Biology Commons, and the Genetics Commons Dartmouth Digital Commons Citation Zeigler, Brandon M.; Sugiyama, Daisuke; Chen, Michael; Guo, Yalin; Downs, K. M.; and Speck, N. A., "The Allantois and Chorion, when Isolated before Circulation or Chorio-Allantoic Fusion, have Hematopoietic Potential" (2006). Open Dartmouth: Published works by Dartmouth faculty. 734. https://digitalcommons.dartmouth.edu/facoa/734 This Article is brought to you for free and open access by the Faculty Work at Dartmouth Digital Commons. It has been accepted for inclusion in Open Dartmouth: Published works by Dartmouth faculty by an authorized administrator of Dartmouth Digital Commons. For more information, please contact [email protected]. Authors Brandon M. Zeigler, Daisuke Sugiyama, Michael Chen, Yalin Guo, K. M. Downs, and N. A. Speck This article is available at Dartmouth Digital Commons: https://digitalcommons.dartmouth.edu/facoa/734 RESEARCH ARTICLE 4183 Development 133, 4183-4192 (2006) doi:10.1242/dev.02596 The allantois and chorion, when isolated before circulation or chorio-allantoic fusion, have hematopoietic potential Brandon M. Zeigler1, Daisuke Sugiyama1,*, Michael Chen1, Yalin Guo1, Karen M. Downs2,† and Nancy A. -

Human Embryologyembryology
HUMANHUMAN EMBRYOLOGYEMBRYOLOGY Department of Histology and Embryology Jilin University ChapterChapter 22 GeneralGeneral EmbryologyEmbryology DevelopmentDevelopment inin FetalFetal PeriodPeriod 8.1 Characteristics of Fetal Period 210 days, from week 9 to delivery. characteristics: maturation of tissues and organs rapid growth of the body During 3-5 month, fetal growth in length is 5cm/M. In last 2 month, weight increases in 700g/M. relative slowdown in growth of the head compared with the rest of the body 8.2 Fetal AGE Fertilization age lasts 266 days, from the moment of fertilization to the day when the fetal is delivered. menstrual age last 280 days, from the first day of the last menstruation before pregnancy to the day when the fetal is delivered. The formula of expected date of delivery: year +1, month -3, day+7. ChapterChapter 22 GeneralGeneral EmbryologyEmbryology FetalFetal membranesmembranes andand placentaplacenta Villous chorion placenta Decidua basalis Umbilical cord Afterbirth/ secundines Fusion of amnion, smooth chorion, Fetal decidua capsularis, membrane decidua parietalis 9.1 Fetal Membranes TheThe fetalfetal membranemembrane includesincludes chorionchorion,, amnion,amnion, yolkyolk sac,sac, allantoisallantois andand umbilicalumbilical cord,cord, originatingoriginating fromfrom blastula.blastula. TheyThey havehave functionsfunctions ofof protection,protection, nutrition,nutrition, respiration,respiration, excretion,excretion, andand producingproducing hormonehormone toto maintainmaintain thethe pregnancy.pregnancy. delivery 1) Chorion: villous and smooth chorion Villus chorionic plate primary villus trophoblast secondary villus extraembryonic tertiary villus mesoderm stem villus Amnion free villus decidua parietalis Free/termin al villus Stem/ancho chorion ring villus Villous chorion Smooth chorion Amniotic cavity Extraembyonic cavity disappears gradually; Amnion is added into chorionic plate; Villous and smooth chorion is formed. -

Study of the Murine Allantois by Allantoic Explants
Developmental Biology 233, 347–364 (2001) doi:10.1006/dbio.2001.0227, available online at http://www.idealibrary.com on Study of the Murine Allantois by Allantoic Explants Karen M. Downs,1 Roselynn Temkin, Shannon Gifford, and Jacalyn McHugh Department of Anatomy, University of Wisconsin–Madison Medical School, 1300 University Avenue, Madison, Wisconsin 53706 The murine allantois will become the umbilical artery and vein of the chorioallantoic placenta. In previous studies, growth and differentiation of the allantois had been elucidated in whole embryos. In this study, the extent to which explanted allantoises grow and differentiate outside of the conceptus was investigated. The explant model was then used to elucidate cell and growth factor requirements in allantoic development. Early headfold-stage murine allantoises were explanted directly onto tissue culture plastic or suspended in test tubes. Explanted allantoises vascularized with distal-to-proximal polarity, they exhibited many of the same signaling factors used by the vitelline and cardiovascular systems, and they contained at least three cell types whose identity, gene expression profiles, topographical associations, and behavior resembled those of intact allantoises. DiI labeling further revealed that isolated allantoises grew and vascularized in the absence of significant cell mingling, thereby supporting a model of mesodermal differentiation in the allantois that is position- and possibly age-dependent. Manipulation of allantoic explants by varying growth media demonstrated that the allantoic endothelial cell lineage, like that of other embryonic vasculatures, is responsive to VEGF164. Although VEGF164 was required for both survival and proliferation of allantoic angioblasts, it was not sufficient to induce appropriate epithelial- ization of these cells.