Published Version
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Host-Pathogen Interactions in Root Infecting Oomycete Species
Host-Pathogen Interactions in Root Infecting Oomycete Species Sara Hadji Mollahossein Faculty of Natural Resources and Agricultural Sciences Department of Forest Mycology and Plant Pathology Uppsala Doctoral Thesis Swedish University of Agricultural Sciences Uppsala 2015 Acta Universitatis Agriculturae Sueciae 2015:9 Cover: Pea plant infected with Phytophthora pisi (left), control plant (right). Sporangia releasing zoospores (left) and oospores (right) in infected root tissue. (Photo: Sara Hosseini) ISSN 1652-6880 ISBN (print version) 978-91-576-8216-1 ISBN (electronic version) 978-91-576-8217-8 © 2015 Sara Hadji Mollahossein, Uppsala Print: SLU Service/Repro, Uppsala 2015 Host-Pathogen Interactions in Root Infecting Oomycete Species Abstract The oomycetes include some of the most devastating pathogens on both cultivated crops and wild plants. In the genus Phytophthora some closely related species have a broad host range, while others are very host specific. The aim of this project was to gain an understanding of the mechanisms underlying the differentiation of a subgroup of root-infecting Phytophthora species and to gain knowledge about the plant immune responses triggered by distantly related oomycetes that adapted to the same legume host. We investigated the zoospore chemotaxis of legume-root infecting Phytophthora species to different isoflavonoid compounds and explored a possible connection to host preference. Our results showed that specific chemotaxis towards host isoflavones is of limited importance in Phytophthora sojae and Phytophthora vignae, while, specific chemotaxis of Phytophthora pisi and Phytophthora niederhauserii indicated an adaptation to their pathogenicity on the host and lack of pathogenicity on non-host plants. The comparative proteomic study of P. pisi and P. -

Genetic Mapping of Resistance to Aphanomyces Root Rot in Alfalfa *E
Genetic mapping of resistance to Aphanomyces root rot in alfalfa15 Samac, D.A.*, Dornbusch, M. R., Bucciarelli, B., Miller, S. S. and Yu, L.-X. USDA-Agricultural Research Service *e-mail: [email protected] KEYWORDS: Aphanomyces euteiches, genotyping by sequencing, NBS-LRR resistance genes Aphanomyces root rot (ARR), caused by the oomycete Aphanomyces euteiches, is one of the most important yield-limiting factors in production of legumes. In Europe, it is the main limiting factor for pea production while in the United States it is one of the most important diseases of alfalfa and pea (Gaulin et al., 2007). The disease causes stunting and death of alfalfa seedlings with complete stand loss occurring in wet and poorly drained soils. Non-lethal damage to seedling roots reduces forage yields and winter survival. The pathogen can infect adult plants during wet periods to cause loss of feeder roots and root nodules, reducing nitrogen fixation, forage yield, and stand life. Varieties with resistance to ARR became widely available in the 1990s; however, failure of resistant varieties identified a second race of the pathogen. Recent reports of failure of varieties with resistance to both race 1 and race 2 suggest that additional pathogenic races are present in alfalfa production fields. The goals of this research were to: (i) determine the prevalence of race 1 and race 2 strains in Minnesota and New York; (ii) identify novel strains of the pathogen; (iii) characterize the race-specific resistance to Aphanomyces root rot; and (iv) map resistance genes in order to facilitate breeding for resistance and to clarify race/resistance gene relationships. -

Development of New Genome-Informed Genotyping Tools for Aphanomyces Astaci
Development of new genome-informed genotyping tools for Aphanomyces astaci Submitted by Diana Minardi to the University of Exeter as a thesis for the degree of Doctor of Philosophy in Biological Sciences In May 2017 This thesis is available for Library use on the understanding that it is copyright material and that no quotation from the thesis may be published without proper acknowledgement. I certify that all material in this thesis which is not my own work has been identified and that no material has previously been submitted and approved for the award of a degree by this or any other University. Signature: ………………………………………………………….. 1 Acknowledgements I would like to sincerely thank my supervisors Dr Mark van der Giezen, Dr David Studholme, and Dr Birgit Oidtmann for their invaluable guidance and support during these 3 years (plus a bit) of PhD, both practically and in the writing of the thesis. I would like to thank Cefas and the University of Exeter for funding the project and for giving me the opportunity to embrace this challenge and adventure. I would like to thank all of the University of Exeter “Biocat” lab and office, with whom I’ve spent most of my coffee time. But especially Mirella (for the laughs and cries) and Simone (for the sweaty runs around campus). I would also like to thank Chiara for her constant support, encouragement, and beer. Finally, I would like to thank Tony, for his patience, and my family, for their “overseas” support: I’ve finally finished “the shrimp” book! 2 Abstract Aphanomyces spp. are water moulds, eukaryotic fungus-like organisms, belonging to the class Oomycota. -

Root Rots in Pulses
CROP DEVELOPMENT CENTRE Root Rots in Pulses Michelle Hubbard, Agriculture and Agri-Food Canada, Swift Current Research Scientist, Pulse Pathology Co-authors: Sabine Banniza, Zakir Hossain, Sherrilyn Phelps, Syama Chatterton, Luke Bainard & Barb Ziesman What is root rot? Caused by micro-organisms Range of pathogens • Fusarium species • Pythium species • Rhizoctonia solani • Aphanomyces euteiches Pea and lentil do not like wet feet High soil moisture can cause • ↓ root and shoot growth • yellowing • ↓ nodulation Stressed plants are more susceptible to infection Normal soil Water- moisture saturated Peas in sterile field soil Root rot is a complex Fusarium species Pythium species Rhizoctonia solani Aphanomyces euteiches And it is complicated! Wider host range True fungi • Fusarium spp. (e.g. solani, • Fusarium spp. avenaceum, acuminatum, graminearum) • Rhizoctonia solani • Rhizoctonia solani • Pythium spp. Fungus-like organisms (oomycetes) Attack only specific plants: • Pythium spp. • Fusarium oxysporum f.sp. pisi, f.sp. ciceris or f. sp. • Aphanomyces lentis, F. virgulifome euteiches • Aphanomyces euteiches • Phytophthora spp. • Phytophthora spp Fusarium Courtesy of S. Chatterton, AAFC Infects many different plants Courtesy of F. Dokken-Bouchard, SMA Aphanomyces Oospores Infects pea and lentil Oospores = resting spores • More vulnerable after they germinate Zoospores: can swim short distances Courtesy of S. Chatterton, AAFC Courtesy of F. Dokken-Bouchard, SMA Aphanomyces Oospores EveryInfects time pea aand plant lentil gets infected,Oospores -
Meeting Program
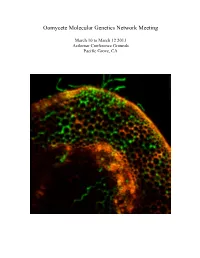
Oomycete Molecular Genetics Network Meeting March 10 to March 12 2013 Asilomar Conference Grounds Pacific Grove, CA Oomycete Molecular Genetics Network Meeting March 10 to March 12, 2013 Asilomar Conference Grounds Pacific Grove, CA The Oomycete Molecular Genetics Research Network was initially funded by an NSF Research Coordination Network grant in 2001. The purpose of our annual meeting is to promote communication and collaboration, and minimize duplication of effort within the worldwide Oomycete molecular genetics community. The oomycete molecular genetics community now numbers well over one hundred labs from across the world, and papers on oomycetes attract a readership that goes well beyond the community itself. The Oomycete Molecular Genetics Conference alternates between USA and Eurasia, and returns this year to Asilomar, California. This year’s meeting will cover some of the latest research on Effector Biology, Genomics and Oomycete evolution and population biology. With over 100 attendees expected, this meeting represents the largest network meeting held in the US. Committee Chairs John McDowell Mark Gijzen Associate Professor Research Scientist Plant Pathology, Physiology and Weed Science Agriculture and Agri-food Canada Virginia Tech, Blacksburg, VA 24061 London ON, Canada NSV4T3 Meeting Logistics Facilities Coordinator Joel Shuman Paul Morris Project Manager Professor, Biological Sciences Plant Pathology, Physiology and Weed Science Bowling Green State University Virginia Tech, Blacksburg, VA24061 Bowling Green, OH 43403 Cover: Confocal image of GFP-expressing Phytophthora sojae at 12h after inoculation of soybean hypocotyls. Image courtesy of Kai-Tao from the lab of Yuanchao Wang, Nanjing Agricultural University, Nanjing, China. Meeting Sponsors This meeting is supported in part by NSF grant MCB-0639226 to Brett Tyler OMGN 2013 Program Location: Talks will be held at Kiln on Sunday Morning and at Fred Farr for all other sessions. -

APHANOMYCES ROOT ROT in PEAS and LENTILS in Western Canada
APHANOMYCES ROOT ROT IN PEAS AND LENTILS in Western Canada albertapulse.com manitobapulse.ca saskpulse.com @albertapulse @mbpulsegrowers @saskpulse ROOT ROTS Root rot in peas and lentils is caused by a complex of diseases that affect the belowground portion of the developing plant, leading to poor performing pulse crops. The organisms that cause the disease are seed- or soil-borne and can infect the plant at any stage. Unfortunately, once root rot has set in, there is nothing that can be done. Understanding the disease, identifying the risks for root rot infection, and thorough planning for prevention are the only current options. Aphanomyces Root Rot DNA testing conducted on root rots in Alberta, Manitoba Aphanomyces root rot is caused by Aphanomyces and Saskatchewan in 2017 (see survey results below), euteiches, a highly specialized pathogen of legumes. show that Aphanomyces is a common cause of root rots While this pathogen has a number of legume host plants, in pulse crops in the Prairies. Aphanomyces can infect peas and lentils are the most susceptible pulse crops to at any time in the growing season and spores persist for infection. Faba beans and sainfoin exhibit good partial PDQ\\HDUVLQWKHVRLOPDNLQJLWWKHPRVWGLI¿FXOWFDXVH (quantitative) resistance to Aphanomyces, and chickpeas of root rot to manage (and therefore the most serious are considered moderately resistant. Soybeans and among the root rot pathogens). While research is being fenugreek are both non-host crops to A. euteiches. undertaken in Alberta, Manitoba and Saskatchewan, there is currently no reliable prevention or cure. Susceptibility of dry beans and alfalfa to Aphanomyces root rot infection varies among the different varieties. -

Trichoderma: a Possible Solution for Aphanomyces ?
Trichoderma: A possible solution for Aphanomyces ? Pratibha Prashar Postdoctoral Fellow Department of Plant Sciences College of Agriculture and Bioresources University of Saskatchewan Trichoderma: A possible solution? • This ascomycete has been established as one of the most acceptable and successful biocontrol agent for a range of soil-borne pathogens. • Across the globe, more than 50 Trichoderma-based products are commercially available as biofungicides, biostimulants, and biological soil amendments (Woo and Lorito,2007). • T. harzianum, T. viride and T. hamatum are some of the most successful species. • There are a few reports for successful control of the other root-rot oomycete pathogen, Pythium, by Trichoderma (Mbarga et al. 2012). • Dandurand et al. (2000) reported synergistic effect of T. harzianum and Brassica napus seed meal for complete suppression of Aphanomyces euteiches root rot in pea. Trichoderma in lentil • Few successful reports for control of soil-borne pathogens particularly Fusarium oxysporum Reference Strain Pathogen Disease Effects Place Garkoti et T.harzianaum+ F.oxysporum Wilt • Reduced disease incidence India al. (2014) P.fluorescens f. sp. lentis • Improved grain yield El-Hassan T.hamatum F.oxysporum Vascular • Delayed infection time UK et al. (Isolate from f. sp. lentis Wilt • Higher pathogen mortality (2013) lentil crop, • Reduced pathogen Syria) colonization Hannan et T.harzianaum + F.oxysporum Foot Rot • Reduced post-emergence Bangladesh al. (2012) Rhizobium + Sclerotium plant death leguminosarum rolfsii • 19/85% higher emergence • Higher plant stand • Up-to 75.56% higher biomass Kashem et T.harzianaum F.oxysporum Foot & • Reduced disease incidence Bangladesh al. (2011) (Native isolate Schlecht Root Rot of 6.9% against 39.0% in from pulse control fields crop) • Improved seed germination, plant stand and seed yield Our Objectives To isolate and screen native as well as other strains of Trichoderma and their secondary metabolites (SMs) for antagonistic potential against Aphanomyces euteiches in lentil. -

Detection of Aphanomyces Euteiches in Field Soil from Northern Idaho by a Wet-Sieving/Baiting Technique
of penetration and movement of fire blight Pathol. 10:178-182. Erwinia amylovora. (Abstr.) Phytopathology bacteria in apple leaf and stem tissue. 23. Sprague, R., and Covey, R. P. 1969. Fungous 74:825. Phytopathology 55:719-723. and bacterial pear diseases of eastern Washing- 30. van der Zwet, T., Bell, R. L., and Stroo, H. 18. Miller, T. D., and Schroth, M. N. 1972. ton. Wash. Agric. Exp. Stn. Circ. 498. F. 1982. Long distance dissemination of Erwinia Monitoring the epiphytic population of Erwinia 24. Thomson, S. V. 1986. The role of the stigma amylovora as resident bacteria in apparently amylovora on pear with a selective medium. in fire blight infections. Phytopathology 76:476- healthy pear budwood. (Abstr.) Phytopathology Phytopathology 62:1175-1182. 482. 72:711. 19. Plurad, S. B., Goodman, R. N., and Enns, W. 25. Thomson, S. V., Schroth, M. N., Reil, W. O., 31. van der Zwet, T., and Keil, H. L. 1972. R. 1967. Factors influencing the efficacy of and Moller, W. J. 1975. Incidence of Erwinia Importance of pear tissue injury to infection by Aphis pomi as a potential vector for Erwinia amylovora in Bartlett pear flowers of different Erwinia amylovora and control with strepto- amylovora. Phytopathology 57:1060-1063. ages. (Abstr.) Proc. Am. Phytopathol. Soc. 2:67. mycin. Can. J. Microbiol. 18:893-900. 20. Roberts, R. G., and Reymond, S. T. 1989. Evalu- 26. University of California. 1965. Do summer oil 32. van der Zwet, T., Thomson, S. V., Covey, R. ation of post-harvest treatments for eradication sprays favor fire blight development in pear P., and Bonn, W. -

Aphanomyces Euteiches Laurent Camborde
Fuctional characterization of different candidate effectors from the root rot oomycete Aphanomyces euteiches Laurent Camborde To cite this version: Laurent Camborde. Fuctional characterization of different candidate effectors from the root rot oomycete Aphanomyces euteiches. Vegetal Biology. Université Paul Sabatier - Toulouse III, 2020. English. NNT : 2020TOU30227. tel-03208760 HAL Id: tel-03208760 https://tel.archives-ouvertes.fr/tel-03208760 Submitted on 26 Apr 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Abstract Oomycetes are eukaryote pathogens able to infect plants and animals. During host interaction, oomycetes secrete various molecules, named effectors, to counteract plant defence and modulate plant immunity. Two different classes of cytoplasmic effectors have been described to date, Crinklers (CRNs) and RxLR proteins. The translocation process allowing the entrance into the host cells is still unclear, and while extended research gave insight into some molecular targets and role during infection, most of effectors have not been characterized. In the root rot pathogen of legumes Aphanomyces euteiches, only the CRNs are present. Based on a previous study reported by our research group, we published an opinion paper focused on the emergence of DNA damaging effectors and their role during infection. -

APHANOMYCES ROOT ROT in PEAS and LENTILS in Western Canada
APHANOMYCES ROOT ROT IN PEAS AND LENTILS in Western Canada albertapulse.com manitobapulse.ca saskpulse.com @albertapulse @mbpulsegrowers @saskpulse ROOT ROTS Root rot in peas and lentils is caused by a complex of diseases that affect the belowground portion of the developing plant, leading to poor performing pulse crops. The organisms that cause the disease are seed- or soil-borne and can infect the plant at any stage. Unfortunately, once root rot has set in, there is nothing that can be done. Understanding the disease, identifying the risks for root rot infection, and thorough planning for prevention are the only current options. Aphanomyces Root Rot DNA testing conducted on root rots in Alberta, Manitoba Aphanomyces root rot is caused by Aphanomyces and Saskatchewan in 2017 (see survey results below), euteiches, a highly specialized pathogen of legumes. show that Aphanomyces is a common cause of root rots While this pathogen has a number of legume host plants, in pulse crops in the Prairies. Aphanomyces can infect peas and lentils are the most susceptible pulse crops to at any time in the growing season and spores persist for infection. Faba beans and sainfoin exhibit good partial PDQ\\HDUVLQWKHVRLOPDNLQJLWWKHPRVWGLI¿FXOWFDXVH (quantitative) resistance to Aphanomyces, and chickpeas of root rot to manage (and therefore the most serious are considered moderately resistant. Soybeans and among the root rot pathogens). While research is being fenugreek are both non-host crops to A. euteiches. undertaken in Alberta, Manitoba and Saskatchewan, there is currently no reliable prevention or cure. Susceptibility of dry beans and alfalfa to Aphanomyces root rot infection varies among the different varieties. -

Evaluation of Soil Bacteria As Bioinoculants for the Control of Field Pea Root Rot Caused by Aphanomyces Euteiches
Evaluation of soil bacteria as bioinoculants for the control of field pea root rot caused by Aphanomyces euteiches A Thesis Submitted to the College of Graduate and Postdoctoral Studies in Partial Fulfilment of the Requirements for the Degree of Master of Science in the Department of Soil Science University of Saskatchewan Saskatoon By Ashebir Tsedeke Godebo © Copyright Ashebir Tsedeke Godebo, March 2019. All rights reserved. PERMISSION TO USE In presenting this thesis in partial fulfilment of the requirement for a post graduate degree from the University of Saskatchewan, I agree that the library of this University may take it freely available for inspection. I further agree that permission for copying of this thesis in any manner, in whole or in part, for scholarly purpose may be granted by the professor or professors who supervised my thesis work or, in their absence, by the Head of the Department or the Dean of the College in which my thesis work is done. It is understood that any copying or publication or use of this thesis part or its parts for financial gain shall not be allowed without my written permission. It is also understood that due consideration shall be given to me and to the University of Saskatchewan in any scholarly use which may be made of any material in my thesis. Request for permission to copy or to make other use of any material in this thesis in whole or part should be addressed to: Head of the Department of Soil Science 51 Campus drive University of Saskatchewan Saskatoon, Saskatchewan S7N 5A8 Canada OR Dean College of Graduate and Postdoctoral Studies University of Saskatchewan 116 Thorvaldson Building, 110 Science Place Saskatoon, Saskatchewan S7N 5C9 Canada i DISCLAIMER Reference in this thesis to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise, does not constitute or imply its endorsement, recommendation, or favoring by the University of Saskatchewan. -

Root Rot in Peas and Lentils
ROOT ROT IN PEAS AND LENTILS in Western Canada Root rot in peas and lentils is caused by a complex of diseases that affect the belowground portion of the developing plant, leading to poor performing pulse crops. The organisms that cause the disease are seed or soil-borne and can infect the plant at any stage. Unfortunately once root rot has set in, there is nothing that can be done. Understanding the disease, identifying the risks for root rot infection, and thorough planning for prevention are the only current options. TABLE OF 2016 Root Rot Survey Results .......................2 CONTENTS Root Rot Symptoms ................................2 Root Rot Pathogens ................................3 Risk Factors for Root Rot ............................4 Root Rot Diagnosis .................................5 Management of Root Rot ...........................6 Research on Root Rot ...............................8 2016 ROOT ROT SURVEY RESULTS Researchers in Alberta, Manitoba, and Saskatchewan have been conducting surveys to assess presence of root rot and levels of severity. Percentage of Fields Testing Positive for Specific Root Rot Pathogens in 2016 Using DNA Testing Number of Aphanomyces Pythium Pythium Fusarium Fusarium Fusarium Fusarium Fusarium Location Crop Fields Tested1 euteiches2 ultimum irregulare avenaceum solani redolens culmorum graminearum Peas 62 61 18 41 88 82 82 23 2 Alberta Lentils 21 29 30 65 91 74 87 39 13 Peas 66 44 33 41 97 94 91 38 27 Saskatchewan Lentils 62 32 33 57 95 70 87 43 22 Manitoba Peas 26 58 28 20 100 88 96 12 4 1 Only fields that showed a disease severity > 3 in the surveys were tested for pathogen composition. 2 These tests are being repeated, results only show fields that had a strong positive reaction to Aphanomyces euteiches.