SWAP Full Document
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
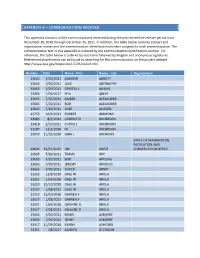
Appendix B – Communications Received
APPENDIX B – COMMUNICATIONS RECEIVED This appendix contains all the communications received during the post-centerline release period from November 18, 2010 through December 31, 2011. In addition, the table below contains contact and organization names and the communication identification number assigned to each communication. The communication text in this appendix is ordered by the communication identification number. For reference, the table below is ordered by last name followed by illegible and anonymous signatures. Referenced attachments can be found by searching for the communication on the project website http://www.bpa.gov/corporate/I-5-EIS/search.cfm. Number Date Name - First Name - Last Organization 13665 7/20/2011 ANDREW ABBOTT 13665 7/20/2011 JACK ABERNATHY 13665 7/20/2011 CRYSTAL L ADAMS 13395 1/29/2011 PHIL AKELY 13667 7/20/2011 AMBER ALEXANDER 13665 7/20/2011 BOB ALEXANDER 13665 7/20/2011 CHEE ALLISON 13755 10/6/2011 ROBERT AMMONS 13683 8/3/2011 CANDICE D ANDERSON 13418 2/10/2011 CURTIS L ANDERSON 13207 12/2/2010 M. ANDERSON 13073 11/22/2010 GINA L ANDREWS STATE OF WASHINGTON, RECREATION AND 13836 12/15/2011 JIM ANEST CONSERVATION OFFICE 13665 7/20/2011 TRAVIS APP 13665 7/20/2011 BOB APPLING 13665 7/20/2011 JEREMY ARIONUS 13665 7/20/2011 CHUCK ARNST 13322 12/8/2010 DALE W AROLA 13321 12/9/2010 DALE W AROLA 13320 12/10/2010 DALE W AROLA 13527 1/28/2011 DALE W AROLA 13320 12/10/2010 DARREN F AROLA 13527 1/28/2011 DARREN F AROLA 13321 12/9/2010 DWAYNE D AROLA 13527 1/28/2011 DWAYNE D AROLA 13665 7/20/2011 BRIAN ASBURRY 13665 -

Older Forests Used by Northern Spotted Owls Functioned As Re Refugia
Older forests used by northern spotted owls functioned as re refugia during large wildres, 1987–2017 Damon B Lesmeister ( [email protected] ) USDA Forest Service Pacic Northwest Research Station https://orcid.org/0000-0003-1102-0122 Raymond J. Davis USDA Forest Service Region 6: USDA Forest Service Pacic Northwest Region Stan G. Sovern USDA Forest Service Pacic Northwest Research Station Zhiqiang Yang USDA Forest Service Rocky Mountain Research Station Research Article Keywords: Northern spotted owl, Strix occidentalis caurina, wildre severity, RdNBR, climate change, re refugia Posted Date: March 12th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-280175/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/27 Abstract Background The northern spotted owl (Strix occidentalis caurina) is an Endangered Species Act-listed subspecies that requires forests with old-growth characteristics for nesting. With climate change, large, severe wildres are expected to be more common and an increasing threat to spotted owl persistence. Understanding re severity patterns related to nesting forest can be valuable for forest management that supports conservation and recovery, especially if nesting forest functions as re refugia (i.e., lower re severity than surrounding landscape). We examined the relationship between re severity and nesting forests in 472 large wildres (> 200 ha) that occurred rangewide during 1987–2017. We mapped re severities (unburned-low, moderate, high) within each re using relative difference normalized burn ratios and quantied differences in severity between pre-re nesting forest (edge and interior) and non-nesting forest. We also quantied these relationships within areas of three re regimes (low severity, very frequent; mixed severity, frequent; high severity, infrequent). -

RCFB April 2021 Page 1 Agenda TUESDAY, April 27 OPENING and MANAGEMENT REPORTS 9:00 A.M
REVISED 4/8/21 Proposed Agenda Recreation and Conservation Funding Board April 27, 2021 Online Meeting ATTENTION: Protecting the public, our partners, and our staff are of the utmost importance. Due to health concerns with the novel coronavirus this meeting will be held online. The public is encouraged to participate online and will be given opportunities to comment, as noted below. If you wish to participate online, please click the link below to register and follow the instructions in advance of the meeting. Technical support for the meeting will be provided by RCO’s board liaison who can be reached at [email protected]. Registration Link: https://zoom.us/webinar/register/WN_JqkQAGCrRSOwbHLmg3a6oA Phone Option: (669)900-6833 - Webinar ID: 967 5491 2108 Location: RCO will also have a public meeting location for members of the public to listen via phone as required by the Open Public Meeting Act, unless this requirement is waived by gubernatorial executive order. In order to enter the building, the public must not exhibit symptoms of the COVID-19 and will be required to comply with current state law around personal protective equipment. RCO staff will meet the public in front of the main entrance to the natural resources building and escort them in. *Additionally, RCO will record this meeting and would be happy to assist you after the meeting to gain access to the information. Order of Presentation: In general, each agenda item will include a short staff presentation and followed by board discussion. The board only makes decisions following the public comment portion of the agenda decision item. -

THIRTY-YEAR CLUB PGION Six U.S.Copst Svicgr
THIRTY-YEAR CLUB PGION Six U.S.cOPST SvIcGr VOL. XVII JUNE -1963 THE HEART OF THE TREE What does he plant who plant. * tree? He plants a friend of sun and sky; He plants the flag of breeea free; A shaft of beauty, towering high. He plants a home to heaven anigh, For song and mother-croon of bird In hushed and happy twilight heard. The treble of heaven's harmony - These things he plants who plants a tree What does he plant who plants a tree? He plants cool shade and tender rain, And seed and bud of days to be, And years that fade and flush again; He plants the glory of the plain; He plants the forest's heritage; The harvest of a coming age; The Joy that unborn eyes shall see - These things he plants who plants a tree. S What does he plant who plants a tree? He plants, in sap and leaf and wood, In love of home and loyalty And far-cast thoughts of civil good. His blessings on the neighborhood Who in the hollow of Hi. hand Holds all the growth of all our land. A nation', growth from sea to sea Stirs in hia heart who plante a tree. Henry- Cuyler Bunner S T I M B E R L I N E S June 1963 VOL. XVII - PUBLISHED ANNUALLY BY R-6 FOREST SERVICE 30-YEAR CLUB Editor - Frank Flack Consulting Editors Publication TH. Burgess Biographies - History....,..,.,.Kirk P. Cecil Obituaries .Leslie L. Colvill Photographs Victor H. Flach Policy J. -

RV Sites in the United States Location Map 110-Mile Park Map 35 Mile
RV sites in the United States This GPS POI file is available here: https://poidirectory.com/poifiles/united_states/accommodation/RV_MH-US.html Location Map 110-Mile Park Map 35 Mile Camp Map 370 Lakeside Park Map 5 Star RV Map 566 Piney Creek Horse Camp Map 7 Oaks RV Park Map 8th and Bridge RV Map A AAA RV Map A and A Mesa Verde RV Map A H Hogue Map A H Stephens Historic Park Map A J Jolly County Park Map A Mountain Top RV Map A-Bar-A RV/CG Map A. W. Jack Morgan County Par Map A.W. Marion State Park Map Abbeville RV Park Map Abbott Map Abbott Creek (Abbott Butte) Map Abilene State Park Map Abita Springs RV Resort (Oce Map Abram Rutt City Park Map Acadia National Parks Map Acadiana Park Map Ace RV Park Map Ackerman Map Ackley Creek Co Park Map Ackley Lake State Park Map Acorn East Map Acorn Valley Map Acorn West Map Ada Lake Map Adam County Fairgrounds Map Adams City CG Map Adams County Regional Park Map Adams Fork Map Page 1 Location Map Adams Grove Map Adelaide Map Adirondack Gateway Campgroun Map Admiralty RV and Resort Map Adolph Thomae Jr. County Par Map Adrian City CG Map Aerie Crag Map Aeroplane Mesa Map Afton Canyon Map Afton Landing Map Agate Beach Map Agnew Meadows Map Agricenter RV Park Map Agua Caliente County Park Map Agua Piedra Map Aguirre Spring Map Ahart Map Ahtanum State Forest Map Aiken State Park Map Aikens Creek West Map Ainsworth State Park Map Airplane Flat Map Airport Flat Map Airport Lake Park Map Airport Park Map Aitkin Co Campground Map Ajax Country Livin' I-49 RV Map Ajo Arena Map Ajo Community Golf Course Map -

Forestry Research Notes
. , WEYERHAEUSER TIMBER Forestry R esearch N otes Tacoma, Washington BETTER TIMBER CROPS THROUGH INDUSTRIAl FOREST MANAGEMENT Fbrestry Research Note Noo 22 January 1960 DOUGLAS - FIR CONE AND SEED INSECT BI OLOGICAL AND CONTROL STUDIES: PROGRESS IN 1958 , 1959 by 'r. 1/ Norman E .. John son and Jack K. Win jum- Foresters in the Pacific Northwest a ~ e g1v1ng increased attention to use of high quality Douglas-fir seed. I nsect damage is one phase of seed investigations being conducted by our forestry resear ch staff at Centralia. The objective is to devel op a sound basis for control of ·the important cone and seed insects of D6uglas-fir. The reporting of pr ogress from these studies should prove of timely inter•est. The present paper on cone and seed insects of Douglas-fir in wester n Washington and Oregon contains sections on: ( 1) a key to seed--damaging larvae commbn i n green Dougl as-fir ·cones in western Washington, ( 2) the p·e,r iod of emergence and attack of the adults of ·these same larvae., ( 3 ) /chemical control tests and ( 4 ) · pre ... cautions on the use of insecticides. Changes in both the scientific and common names of the insects dealt with in this paper has resulted in some confusion. The following l ist of "Names usedr·r with the "Explanatory notes" is included to give the various synonyms with references where the name was originally used. The authors ., Forest Entomol ogist and Technol ogi·st, r 'espectively, express appreciation for fine cboperation that facilitated these studies: Chemical companies for ~upplying insecticides to test; members of our forestry research staff, D~. -

Fire Management Today Volume 62(3), the Article “Thirtymile Fire: Fire Behavior and Management Response” by Hutch Brown Made Some Incor- Rect Statements
Fire today ManagementVolume 65 • No. 1 • Winter 2005 LLARGEARGE FFIRESIRES OFOF 2002—P2002—PARTART 11 United States Department of Agriculture Forest Service CORRECTIONS ON THIRTYMILE In Fire Management Today volume 62(3), the article “Thirtymile Fire: Fire Behavior and Management Response” by Hutch Brown made some incor- rect statements: • p. 26, col. 1, para. 2: The statement that the fire management officers (FMOs) on Thirtymile “had no recent experience with initial and extend- ed attack” is false. In fact, the FMOs had recent experience on dozens of fires with various initial- and extended-attack resources, including smokejumpers, helirappelers, contractors, and ground resources. Those involved in formulating strategy and tactics on Thirtymile, including the FMOs, incident commander, and hotshot supervisor and foreman, had decades of fireline experience among them. • p. 26, col. 3, para. 3: The statement that engines arriving on the fire drove past the fire crew “[w]ithout checking in with the IC” has been proven to be false. The assigned incident commander remembers that engine 701 did check in at the lunch spot. • p. 29, col. 1, para. 1: The reference to media stories as “independent investigations” is misleading in a journal for the international wildland fire community. Newsprint journalism should not be confused with a formal accident investigation following a tragedy fire. We apologize to readers for the inaccuracies in the article and especially to those who might have been affected by them. Fire Management Today is published by the Forest Service of the U.S. Department of Agriculture, Washington, DC. The Secretary of Agriculture has determined that the publication of this periodical is necessary in the transaction of the pub- lic business required by law of this Department. -

TRI-POWER Is to Enlighten Fellow Enthusiasts by Educating, Inspiring and Informing
FEBRUARYFEBRUARY 20142014 TRI-RI-POWEROWER Quadra Paws Fun Run 2013-Pages...10-11 Golf ball toss Wheel spin for points Cline Butte Work Party - March 22nd -2013....Pages 4-5 Lava rocks form rims and they present a perfect playground These Class II users belong to various clubs in Oregon PAID PERMIT #88 US POSTAGE GRESHAM, OR PRESORTED STD. www.pnw4wda.org 4-Wheelin’ News FEBRUARY 2014 1 PNWPNW4WDAWDA TRI-RI-POWEROWER www.pnw4wda.org / 800-537-7845 Volume 43 Number 2 February 2014 The mission of TRI-POWER is to enlighten fellow enthusiasts by educating, inspiring and informing Editor/Publisher: Ron McDonald / 360-901-8376 * Email: [email protected] 3902 NE 61st Ave, Vancouver, WA 98661 * Messages (360) 695-0595 All Rights Reserved - Printed in the USA TRI-POWER is published monthly for the Pacific Northwest 4 Wheel Drive Association. Photographs are edited and prepared with Adobe Photoshop.Layout is performed in Adobe InDesign. In house ads are created with Adobe Illustrator. Prepress product is converted with Adobe Acrobat Professional. TRI-POWER makes no representation or warranty, express or implied, with respect to the completeness, accuracy or utility of these materials or any information or opinion contained herein. Any use or reliance on the information or opinion is at the risk of the user, and TRI-POWER shall not be liable for any damage or injury incurred by any person arising out of the completeness, accuracy or utility of any information or opinion contained in these materials. These materials are not to be construed as an endorsement of any product or company, nor as the adoption or promulgation of any guidelines, standards, or recommendation. -

HRCCWPP 2013 Draft1.1
Table of Contents Introduction ..............................................................................................................7 Vision ..............................................................................................................8 Goals ...............................................................................................................9 Purpose ............................................................................................................9 Who 9 What 10 Where ............................................................................................................10 Why 10 When .............................................................................................................10 How 11 Organization ..................................................................................................11 Funding .........................................................................................................12 Impetus and Planning Steps ...................................................................................14 Policies and Tools .........................................................................................14 Defining the Wildland/Urban Interface ........................................................16 Planning Steps ...............................................................................................17 The 2005 Hood River CWPP Planning Process ...........................................18 Hood River CWPP Update Process ..............................................................18 -

2.2 State Risk Assessment
Chapter 2: RISK ASSESSMENT | State Risk Assessment | Cultural Resources Summary » Wildfire 2.2 State Risk Assessment Requirement: 44 CFR §201.4(c)(2)(i): The risk assessment shall include… (i) An overview of the type and location of all natural hazards that can affect the State, including information on previous occurrences of hazard events, as well as the probability of future hazard events, using maps where appropriate; The spatial distribution of the facilities within hazard zones is not easily viewed on a statewide map. Therefore, maps depicting hazard zones and facilities within those zones have only been created at the regional scale. Those maps can be found in section 2.3, Regional Risk Assessments. Oregon Natural Hazards Mitigation Plan | September 2020 163 Chapter 2: RISK ASSESSMENT | State Risk Assessment | Coastal Hazards Summary » Wildfire 2.2.1 Coastal Hazards The Pacific Northwest (PNW) coast of Figure 2-29. Erosion at The Capes Oregon is without doubt one of the most Condominiums, Oceanside, Oregon dynamic coastal landscapes in North America, evident by its long sandy beaches, sheer coastal cliffs, dramatic headlands and vistas, and ultimately the power of the Pacific Ocean that serves to erode and change the shape of the coast. It is these qualities along with its various natural resources that have drawn people to live along its narrow shores. However, coastal communities are increasingly under threat from a variety of natural hazards that all come together along the coastal strip. These include wave-induced coastal erosion (both short and long term), wave runup and overtopping (wave-induced flood hazards), inundation Notes: The Capes, a multi-million dollar condominium of homes by wind-blown sand, coastal complex constructed on an old Holocene dune field landslides, earthquakes, and potentially adjacent to Oceanside. -

Spring/Fall 2001 Forestforest Historyhistory Todaytoday Message from the President by Understanding Our Past, We Shape Our Future
A PUBLICATION OF THE FOREST HISTORY SOCIETY SPRING/FALL 2001 ForestForest HistoryHistory TodayToday MESSAGE FROM THE PRESIDENT By understanding our past, we shape our future STEVEN ANDERSON ne of the most poignant lessons “It’s interesting that before the United from history is that the prosperi- Nations and before the World Trade ty of civilizations is based both Center, Gifford Pinchot dreamed of a Oon the availability and use of natural World Conservation Conference, which resources and on the social interactions finally resulted in the “Lake Success between people and nations. In many ways Conference.” His wife termed the confer- what man does to the land, man does to man. Gifford ence “tragic” (FHT Spring 1999, p. 41), and she describes Pinchot recognized this as he spent many of his last years for- men of little faith and many fears. Two world wars and mulating plans for an international conference on conserva- many revolutions marked the first half of the 20th century. tion that would set a foundation for permanent peace. Pinchot’s goal of a permanent peace and prudent utiliza- The world must and is responding to the attacks of tion of natural resources for the welfare of all nations has September 11, 2001, events that will be considered among the still not arrived. From images of Afghanistan seen on tele- worst atrocities in American history. It is understandable that vision lately it looks like a pretty desolate place. Pinchot’s the initial response was focused on ensuring immediate safety. vision of conservation and world policy were certainly rel- Yet, reflection to situate the crisis in a historical context also evant during his time. -

Climate Change Vulnerability and Adaptation in the Columbia River Gorge, Mount Hood National Forest, and Willamette National Forest
Climate Change Vulnerability and Adaptation in the Columbia River Gorge, Mount Hood National Forest, and Willamette National Forest Editors Jessica E. Halofsky is the director of the Northwest Climate Hub and Western Wildland Environmental Threat Assessment Center, Pacific Northwest Research Station, 3625 93rd Avenue SW, Olympia, WA 98512; David L. Peterson is a professor, University of Washington, College of the Environment, School of Environmental and Forest Sciences, Box 352100, Seattle, WA 98195-2100; Rebecca A. Gravenmier is a science coordinator and regional climate change coordinator, U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station and Pacific Northwest Region, 1220 SW 3rd Avenue, Portland, OR 97204. 1 Climate Change Vulnerability and Adaptation in Columbia River Gorge, Mount Hood National Forest, and Willamette National Forest Jessica E. Halofsky, David L. Peterson, and Rebecca A. Gravenmier Editors U.S. Department of Agriculture, Forest Service Pacific Northwest Research Station Portland, Oregon General Technical Report PNW-GTR-xxx Month year 2 Abstract Halofsky, Jessica E.; Peterson, David L.; Gravenmier, Rebecca A., eds. 2020. Climate change vulnerability and adaptation in the Columbia River Gorge, Mount Hood National Forest, and Willamette National Forest. Gen. Tech. Rep. PNW-GTR-xxx. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. Xxx p. A science-management partnership was developed to identify climate change issues relevant for resource management in Columbia River Gorge National Scenic Area, Mount Hood National Forest, and Willamette National Forest (abbreviated as CMW Adaptation Partnership, or CMWAP). This partnership assessed the vulnerability of natural resources to climate change, and developed adaptation options that minimize negative impacts of climate change and facilitate transition of ecosystems and organizations to a warmer climate.