Traditional Eastern Spanish Varieties of Tomato
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Denominació Dels Municipis Valencians
DENOMINACIÓ DELS MUNICIPIS VALENCIANS NOM VALENCIÀ NOM OFICIAL GENTILICI PRED. LING. Ademús Ademuz ademusser, ademussera castellà Ador Ador adorer, adorera valencià Agost Agost agoster, agostera valencià Agres Agres agresà, agresana valencià Agullent Agullent agullentí, agullentina valencià Aielo de Malferit Aielo de Malferit aieloner, aielonera valencià Aielo de Rugat Aielo de Rugat aielí, aielina valencià Aigües Aigües aigüer, aigüera valencià Aín Aín aïnenc, aïnenca valencià Aiòder Ayódar aiodà, aiodana aioder, aiodera castellà Aiora Ayora aiorí, aiorina castellà Alacant Alacant / Alicante alacantí, alacantina valencià Alaquàs Alaquàs alaquaser, alaquasera valencià Albaida Albaida albaidí, albaidina valencià Albal Albal albalenc, albalenca valencià Albalat de la Ribera Albalat de la Ribera albalatenc, albalatenca valencià Albalat dels Sorells Albalat dels Sorells albalatà, albalatana valencià Albalat dels Tarongers Albalat dels Tarongers albalatenc, albalatenca valencià Albatera Albatera albaterà, albaterana castellà alberiquer, alberiquera Alberic Alberic valencià alberiqueny, alberiquenya Albocàsser Albocàsser albocassí, albocassina valencià Alboraia Alboraya alboraier, alboraiera valencià Alboraig Alborache alborager, alboragera castellà Albuixec Albuixech albuixequer, albuixequera valencià Alcalà de Xivert Alcalà de Xivert xivertí, xivertina valencià Alcalalí Alcalalí alcalalí, alcalalina valencià Alcàntera de Xúquer Alcàntera de Xúquer alcanterí, alcanterina valencià Alcàsser Alcàsser alcasser, alcassera valencià Alcoi Alcoi -

Venta Del Moro
Proyecto de rehabilitación de la cantera de “La Serratilla” Anejos a la memoria en el t.m de Venta del Moro ÍNDICE Anejo Número 1: Estudio climático 3-10 1-.Elección de observatorios 3 2-. Análisis de los datos climáticos 4-7 3-. Índices y gráficos climáticos 8-10 Anejo Número 2: Estudio socioeconómico 11-21 1-. Estudio de la población 11-15 2-. Estudio del mercado de trabajo 16-19 3-. Diagnóstico de sostenibilidad 20,21 Anejo Número 3: Movimiento de tierras 22-24 1-. Introducción 22 2-. Rellenado 22 3-. Aporte de tierra vegetal 23 4-. Maquinaria 24 Anejo Número 4: Vegetación 25,46 1-. Evolución de la vegetación a lo largo de la historia 25,26 2-. Rasgos florísticos por paisajes vegetales 27-31 3-. Especies de la repoblación 32-46 Proyecto fin de carrera 1 Adrián Moya García Proyecto de rehabilitación de la cantera de “La Serratilla” Anejos a la memoria en el t.m de Venta del Moro Anejo Número 5: Fauna 47-88 1-. Fauna asociada a los distintos ecosistemas 47-51 2-. Clasificación de la fauna del Término 52-88 Anejo Número 6: Estudio de impacto ambiental 89-94 1-. Introducción 90 2-. Identificación de los impactos producidos por la explotación 90-93 3-. Medidas correctoras 93,94 Anejo Número 7: Estudio de seguridad y salud 95-113 1-. Objetivo del presente estudio de seguridad y salud 96 2-. Identificación de la obra 97 3-. Relación resumida de los trabajos a realizar 98 4-. Fases de obra con identificación de riesgos 99 5-. -

17 Juli 1984 Houdende Algemene Voorschriften Inzake De Toekenning Van De Produktiesteun Voor Olijfolie En De in Bijlage II Van Verordening (EEG) Nr
Nr. L 122/64 Publikatieblad van de Europese Gemeenschappen 12. 5. 88 VERORDENING (EEG) Nr. 1309/88 VAN DE COMMISSIE van 11 mei 1988 tot wijziging van Verordening (EEG) nr. 2502/87 houdende vaststelling van de opbrengst aan olijven en aan olie voor het verkoopseizoen 1986/1987 DE COMMISSIE VAN DE EUROPESE geerd, gezien het feit dat de begunstigden de produktie GEMEENSCHAPPEN, steun nog , niet hebben kunnen ontvangen ; Gelet op het Verdrag tot oprichting van de Europese Overwegende dat de in deze verordening vervatte maatre Economische Gemeenschap, gelen in overeenstemming zijn met het advies van het Gelet op Verordening nr. 136/66/EEG van de Raad van Comité van beheer voor oliën en vetten. 22 september 1966 houdende de totstandbrenging van een gemeenschappelijke ordening der markten in de sector oliën en vetten ('), laatstelijk gewijzigd bij Verorde HEEFT DE VOLGENDE VERORDENING ning (EEG) nr. 1098/88 (2), en met name op artikel 5, lid VASTGESTELD : 5, Gelet op Verordening (EEG) nr. 2261 /84 van de Raad van Artikel 1 17 juli 1984 houdende algemene voorschriften inzake de toekenning van de produktiesteun voor olijfolie en de _ In bijlage II van Verordening (EEG) nr. 2502/87 worden steun aan de producentenorganisaties (3), laatstelijk gewij de gegevens betreffende de autonome gemeenschappen zigd bij Verordening (EEG) nr. 892/88 (4), en met name Andalusië en Valencia vervangen door de gegevens die op artikel 19, zijn opgenomen in de bijlage van deze verordening. Overwegende dat bij Verordening (EEG) nr. 2502/87 van Artikel 2 de Commissie (*), gewijzigd bij Verordening (EEG) nr. 370/88 (*), de opbrengst aan olijven en aan olie is vastge Deze verordening treedt in werking op de dag van haar steld voor de homogene produktiegebieden ; dat in bijlage bekendmaking in het Publikatieblad van de Europese II van die verordening vergissingen zijn geconstateerd Gemeenschappen. -

MANUEL | Toponímia Dels Pobles Valencians
MANUEL L A R IBERA A LTA AJUNTAMENT DE MANUEL ACADÈMIA VALENCIANA DE LA LLENGUA SECCIÓ D’ONOMÀSTICA COORDINACIÓ I GESTIÓ Unitat de Recursos Lingüisticotècnics RECULL I TEXT Gonçal Benavent Carbonell Facund Blasco Martínez Pau Martínez Gómez GRAFISME Esperança Martínez Molina © Acadèmia Valenciana de la Llengua Col·lecció: Onomàstica Sèrie: Toponímia dels Pobles Valencians Manuel, 219 Editat per: Publicacions de l’Acadèmia Valenciana de la Llengua Av. de la Constitució, 284 46019 València Tel.: 96 387 40 23 Adreça electrònica: [email protected] Amb la col·laboració de l’Ajuntament de Manuel ISBN: 978-84-482-6226-6 Depòsit legal: V-851-2018 Impressió: www.avl.gva.es MANUEL El municipi de Manuel se situa en la part sud de la comarca de la Ribera Alta. El terme municipal, amb una extensió de sis quilòmetres quadrats, limita al nord amb Sant Joanet; a l’oest, amb Senyera i Vilanova de Castelló; a l’est, amb l’Ènova i la Pobla Llarga, i al sud, amb Xàtiva. L’any 2017 Manuel té 2.448 habitants. El 82 % de la població parla valencià i el 100 % afirma que l’entén, segons les dades del Cens de Població i Vivendes de l’any 2011. El riu d’Albaida travessa el terme de Manuel cap a la ribera del Xúquer, del qual és afluent. La part nord del terme és plana i d’horta, constituïda bàsicament per tarongers. En el sud, estenent-se cap a l’est fins a l’Ènova i Barxeta, es troba la serra de Valiente, amb una gran diversitat de plantes aromàtiques. -

Verification of Vulnerable Zones Identified Under the Nitrate
CONTENTS 1 INTRODUCTION 1 1.1 OVERVIEW OF THE QUALITY OF CONTINENTAL WATERS 1 1.2 PROBLEMS ENCOUNTERED DURING THE INVESTIGATIONS 2 2 PROCEDURES FOR DESIGNATING SENSITIVE AREAS AND VULNERABLE ZONES AND APPLICATION OF CRITERIA ESTABLISHED IN DIRECTIVES 5 2.1 INTRODUCTION 5 2.2 PROCEDURES FOR DESIGNATING SENSITIVE AND LESS SENSITIVE AREAS (DIRECTIVE 91/271/EEC) 8 2.3 PROCEDURES FOR DESIGNATING VULNERABLE ZONES (DIRECTIVE 91/676/EEC). 14 2.4 ANALYSIS OF THE APPLICATION OF CRITERIA FOR DESIGNATION OF SITES UNDER DIRECTIVES 91/271/EEC AND 91/676/EEC. 20 3 CONCLUSIONS REGARDING THE RELATIONSHIP BETWEEN VULNERABLE ZONES AND SENSITIVE AREAS IN SPAIN, TAKING INTO ACCOUNT COMMON STANDARDS. 30 1 INTRODUCTION 1.1 OVERVIEW OF THE QUALITY OF CONTINENTAL WATERS At the end of 1998, the national Ministry of the Environment completed the “Libro Blanco del Agua en España”, or (White Paper for Water in Spain). This document was designed as a tool for assessing and guiding water management in Spain. Although Spain’s future water policy is outlined in the conclusions, the points mentioned refer only to quantitative aspects of water resources without considering the issue of quality. Nonetheless, the issue of river, reservoir, lake and groundwater pollution is taken into account in the assessment section. Amongst the problems related to water quality, the White Paper notes, among others, problems related with Directives 91/271/EEC and 91/676/EEC. A brief summary of the main issues addressed is provided below. Surface water and diffuse pollution. In general, diffuse pollution produced by agricultural and livestock raising activities (e.g. -

Calendarios De Guardia 2021 Juzgados De Instrucción
CALENDARIOS DE GUARDIA 2021 JUZGADOS DE INSTRUCCIÓN PROVINCIA DE VALENCIA ÍNDICE DECANATO DE VALENCIA PÁG. Juzgados de Instrucción 40 Juzgados de Violencia sobre la Mujer 43 Juzgados de Menores 45 OTROS PARTIDOS JUDICIALES Decanato de Alzira 47 Decanato de Carlet 49 Decanato de Catarroja 51 Decanato de Gandía 53 Decanato de Llíria 55 38 Decanato de Massamagrell 57 Decanato de Mislata 59 Decanato de Moncada 61 Decanato de Ontinyent 63 Decanato de Paterna 65 Decanato de Picassent 67 Decanato de Quart de Poblet 69 Decanato de Requena 71 Decanato de Sagunto 73 Decanato de Sueca 75 Decanato de Torrent 77 Decanato de Xàtiva 79 39 JUZGADO DECANO EXCLUSIVO DE VALENCIA CALENDARIO GUARDIAS INSTRUCCIÓN - PRIMER SEMESTRE – 2021 ENERO FEBRERO MARZO ABRIL MAYO JUNIO Día Det. Ind. Del. Día Det Ind Del. Día Det Ind. Del. Día Det. Ind. Del. Día Det. Ind. Del. Día Det. Ind. Del. Lev. Lev. Lev. Lev. Lev. Lev. L 1 17 16 8 1 4 3 17 M 2 18 17 9 2 5 4 18 1 17 15 8 Mi 3 19 18 11 3 6 5 19 2 18 17 9 J 4 20 19 12 4 7 6 20 1 16 15 7 3 19 18 10 V 1 5 4 5 21 20 13 5 8 7 21 2 17 16 4 20 19 11 S 2 6 5 6 1 21 6 9 8 3 18 17 1 5 4 5 21 20 D 3 7 6 7 2 1 7 10 9 4 19 18 2 6 5 6 1 21 L 4 8 7 21 8 3 2 16 8 11 10 3 5 20 19 3 7 6 20 7 2 1 14 M 5 10 8 1 9 4 3 17 9 13 11 4 6 21 20 12 4 8 7 21 8 3 2 15 Mi 6 11 10 10 5 4 18 10 14 13 5 7 1 21 14 5 9 8 1 9 4 3 17 J 7 12 11 3 11 6 5 19 11 15 14 6 8 2 1 15 6 10 9 2 10 5 4 18 V 8 13 12 4 12 7 6 20 12 16 15 7 9 3 2 7 11 10 3 11 6 5 19 S 9 14 13 13 8 7 13 17 16 10 4 3 8 12 11 12 7 6 D 10 15 14 14 9 8 14 18 17 11 5 4 9 13 12 13 8 7 L 11 -

El Regadío Tradicional De La Ribera Alta Del Xúquer Capítulo 1
II EL REGADÍO TRADICIONAL DE LA RIBERA ALTA DEL XÚQUER CAPÍTULO 1 EL REGADÍO EN LA RIBERA ALTA DEL Como singularidad estructural, el Xúquer no ocupa el punto más bajo de su valle, de este sector del gran llano de inundación que es XÚQUER. EL MARGEN DERECHO la Ribera, sino que igual que hemos visto en la Ribera Baixa discurre a mayor altitud que las tierras inmediatas. Ello se debe a la mayor Antonio Furió Diego sedimentación de materiales en el propio cauce del río o en sus márge- Luis Pablo Martínez nes que en las tierras inundables, ya que son más frecuentes los momentos en que el río va lleno que los momentos en que se desborda. El llano de Departament d'Història Medieval inundación adquiere así una forma convexa en la que el Xúquer ocupa la Universitat de València parte más alta y el barranco de Barxeta, que discurre en buena medida en paralelo hasta converger con él, la más honda. En dos de los cortes Dos ríos, un barranco y una calzada transversales practicados por Vicent Ferrer, la superficie baja de los 24 La Ribera del Xúquer, lo hemos dicho ya en otro capítulo de m y 27 m sobre el nivel del mar, al lado del río, a los 19 y 20, respectivamente, este mismo libro, es a la vez una y diversa. Una unidad física, determinada junto al barranco. Es en esta superficie convexa, entre el Xúquer y el por el relieve y la hidrografía, y una unidad humana, conformada por Barxeta, con una pendiente de entre cinco y siete metros, donde se ha siglos de historia compartida. -

Los Robos En Los Hogares De La Comunidad Valenciana Robos En Viviendas De La Comunidad Valenciana: Dónde Y Cuánto Se Roba
Los robos en los hogares de la Comunidad Valenciana Robos en viviendas de la Comunidad Valenciana: dónde y cuánto se roba Los datos e informaciones contenidos en este informe son propiedad de UNESPA. El propietario cede su uso libre por parte de las entidades aseguradoras miembros de la Asociación, así como los medios de comunicación. Cualquier otro agente que desee reproducir la totalidad o parte de la información aquí contenida debe para ello de contar con la autorización expresa del propietario. 1 Índice INTRODUCCIÓN DEL ESTUDIO 4 LOS ROBOS EN HOGARES VALENCIANOS 5 ESTACIONALIDAD DE LOS ROBOS 5 DISTRIBUCIÓN TERRITORIAL DE LOS ROBOS 9 LA IMPORTANCIA DE LA FRECUENCIA 12 GRAVEDAD DE LOS ROBOS: CUÁNTO SE LLEVAN 16 ANEXO DE TABLAS PROVINCIALES 22 ALICANTE 22 CASTELLÓN 27 VALENCIA 30 ROBOS EN LAS VIVIENDAS EN LA CIUDAD DE VALENCIA 37 INTRODUCCIÓN Y METODOLOGÍA 37 ESTACIONALIDAD DE LOS ROBOS DE VALENCIA 37 DISTRIBUCIÓN DE LOS ROBOS POR DISTRITOS DE VALENCIA 39 FRECUENCIA DE ROBOS EN LOS DISTRITOS DE VALENCIA 40 GRAVEDAD DE LOS ROBOS: CUÁNTO SE LLEVAN EN CADA DISTRITO 41 Índice de gráficos Ilustración 1: Estacionalidad por meses de los robos en la Comunidad Valenciana.__________________ 7 Ilustración 2: Estacionalidad por día de la semana de los robos en Andalucía. _____________________ 8 Ilustración 3: Estacionalidad de los robos por meses en las viviendas de Valencia. _________________ 38 Ilustración 4: Estacionalidad de los robos por días en las viviendas de Valencia. ___________________ 38 2 Índice de tablas Tabla 1: Las fechas con más robos estimados en la Comunidad Valenciana. _______________________ 5 Tabla 2: Estacionalidad de los robos en la Comunidad Valenciana, por provincias. -
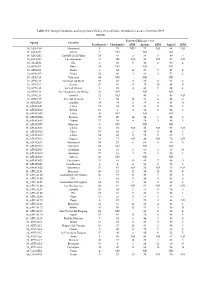
Table S1. Sample Locations and Expected Efficacy of Acaricides Recorded in Assays from the 2018 Season
Table S1. Sample locations and expected efficacy of acaricides recorded in assays from the 2018 season. Expected Efficacy 1 (%) Apiary Location Pyrethroids 2 Checkmite+ SEM 3 Apitraz SEM Amicel SEM 18_ADSAV6 Monserrat 3 73 ND 4 77 ND 94 ND 18_AIXAM1 Altura 3 ND ND ND 18_AIXAM2 Castelló de la Plana 18 69 5 56 5 80 6 18_AIXAM4 Les Alqueries 11 100 ND 93 ND 93 ND 18_ALAPI3 Elx 0 83 5 80 2 90 4 18_APAC01 Betxí 93 ND ND ND 18_APAC02 Onda 0 64 11 72 7 87 4 18_APAC05 Onda 32 46 4 81 4 75 2 18_APAC10 Vila-real 89 ND ND ND 18_APAC11 Sant Joan de Moró 50 47 8 58 2 51 3 18_APAC12 Costur 87 41 12 25 10 64 8 18_APAC13 La Vall d’Uixó 3 72 4 81 7 82 4 18_APAC14 Sta. Magdalena de Pulpis 33 ND ND ND 18_APAC16 Borriol 72 ND 80 6 85 ND 18_APAC17 Ares del Maestre 5 56 12 81 6 86 6 18_APIADS0 Caudete 34 34 2 78 4 81 11 18_APIADS01 Llíria 78 38 14 83 11 90 5 18_APIADS02 Bétera 65 5 4 64 9 94 2 18_APIADS03 Llíria 40 ND ND ND 18_APIADS04 Barxeta 77 40 14 94 3 89 6 18_APIADS05 Cheste 75 34 4 54 3 88 3 18_APIADS07 Manises 0 ND ND ND 18_APIADS11 Chiva 51 72 ND 86 ND 97 ND 18_APIADS12 Chiva 76 41 3 80 4 84 2 18_APIADS13 Llíria 59 60 2 72 0 65 7 18_APIADS14 Sagunt 13 71 ND 89 ND 79 ND 18_APIADS15 Monserrat 55 25 6 66 4 68 5 18_APIADS16 Montroy 22 ND ND ND 18_APIADS17 Gestalgar 65 32 2 65 5 63 10 18_APIADS18 Gestalgar 77 19 4 72 5 79 1 18_APIADS19 Yátova 10 ND ND ND 18_APIADS20 Carcaixent 53 49 10 83 7 89 11 18_APIADS21 Guadassuar 0 4 4 59 2 73 6 18_APIADS22 Monserrat 37 37 ND 70 ND 53 ND 18_APICAL01 Beniarrés 65 12 12 80 12 92 9 18_APICAL02 Guardamar del Segura 60 -

CONTROL MADURACIÓN 14 Histórico Extratempranas 06-10-14
Servicio de Desarrollo Tecnológico SEGUIMIENTO CONTROL DE MADURACIÓN DE CÍTRICOS Campaña 2014-2015 ÍNDICE DE COLOR Hunter (IC=1000a/Lb) VARIEDAD PATRÓN PROVINCIA COMARCA MUNICIPIO 01/09/2014 08/09/2014 15/09/2014 22/09/2014 29/09/2014 06/10/2014 OBSERVACIONES IWASAKI C. CARRIZO VALENCIA LA SAFOR TAVERNES DE LA VALLDIGNA -17,28 RECOLECTADA IWASAKI C. CARRIZO VALENCIA LA RIBERA ALTA CARCAIXENT -12,30 -12,53 -8,24 RECOLECTADA IWASAKI C. CARRIZO VALENCIA CAMP DE TÚRIA LLÍRIA -14,02 -12,43 -10,50 -6,87 RECOLECTADA IWASAKI C. CARRIZO CASTELLÓN LA PLANA BAIXA VILAVELLA -18,50 -11,71 -13,46 RECOLECTADA OKITSU C. CARRIZO VALENCIA LA RIBERA ALTA CARCAIXENT -19,51 -20,59 -16,94 -18,75 -13,97 RECOLECTADA OKITSU C. CARRIZO VALENCIA CAMP DE TÚRIA LLÍRIA -17,50 -15,92 -13,46 -9,32 -3,30 BASOL C. CARRIZO VALENCIA LA RIBERA ALTA CARCAIXENT -9,96 -10,84 -10,40 2,89 RECOLECTADA BASOL CITRUMELO CASTELLÓN LA PLANA BAIXA BETXÍ -19,76 -16,63 -11,66 RECOLECTADA BASOL C. CARRIZO CASTELLÓN LA PLANA ALTA CASTELLÓ -19,59 -16,92 -5,76 RECOLECTADA CLEMENSOON C. CARRIZO VALENCIA LA RIBERA ALTA CARCAIXENT -18,48 -15,29 -13,83 -7,04 6,31 RECOLECTADA CLEMENRUBÍ C. CARRIZO VALENCIA LA SAFOR OLIVA (ELCA) -14,16 RECOLECTADA CLEMENRUBÍ C. MACROPHYLLA VALENCIA LA SAFOR OLIVA (EL SALT) -12,17 RECOLECTADA CLEMENRUBÍ C. CARRIZO VALENCIA LA SAFOR OLIVA (EL SALT) -14,43 RECOLECTADA CLEMENRUBÍ C. CARRIZO VALENCIA LA RIBERA ALTA CARCAIXENT -18,12 -19,39 -17,10 -8,92 1,48 RECOLECTADA CLEMENRUBÍ C. -

Aportaciones a La Flora De Carcaixent (Valencia) Y Áreas Limítrofes
Flora Montiberica 37: 68-74 (XII-2007). ISSN 1138-5952 APORTACIONES A LA FLORA DE CARCAIXENT (VALENCIA) Y ÁREAS LIMÍTROFES Pablo VERA GARCÍA1*, S. Ignacio ENCABO FOS1, Juan S. MONRÓS GONZÁLEZ1, Emilio BARBA CAMPOS1 & Eduardo J. BELDA PÉREZ2 1 Instituto Cavanilles de Biodiversidad y Biología Evolutiva. Universidad de Valencia. Apartado Oficial 22085. 46071 Valencia. 2 E.P.S. de Gandía, Departamento de Ciencia Animal. Universidad Politécnica de Valencia. Crta. Nazaret-Oliva, s/n. 46730 Gandía (Valencia). * [email protected] RESUMEN: Se comenta la presencia y localización de varios taxones raros o novedosos para áreas próximas al municipio de Carcaixent (La Ribera Alta, Valencia), entre las que destacan las poco conocidas Ononis diffusa Ten. y Serapias parviflora Parl. Se discute también el rango taxonómico de algunos ejemplares recolectados del género Delphinium. Palabras clave: flora, distribu- ción, corología, Delphinium, Serapias, Ononis diffusa, Carcaixent, Valencia. SUMMARY: Presence and location from several rare taxa or regional no- velties from near areas of Carcaixent (La Ribera Alta, SE Valencia) are here commented, pointing out Ononis diffusa Ten. and Serapias parviflora Parl. Taxonomic rank of some collected Delphinium is also discussed. Key words: flora, distribution, chorology, Delphinium, Serapias, Ononis diffusa, Carcaixent, Valencia. INTRODUCCIÓN el Río Júcar su límite administrativo al O (Fig. 1). Desde el punto de vista biogeo- Con motivo de la realización del ca- gráfico, el término se encuentra situado en tálogo florístico del término municipal de el límite septentrional de los territorios del Carcaixent (La Ribera Alta, Valencia), subsector Alcoyano-Diánico (sector Seta- durante los años 2006 y 2007 se llevaron bense, provincia Catalano-Valenciano- a cabo un gran número de jornadas botá- Provenzal) (De la TORRE & al., 1996). -

Horario Y Mapa De La Ruta 265 De Autobús
Horario y mapa de la línea 265 de autobús 265 265a Valencia Ver En Modo Sitio Web La línea 265 de autobús (265a Valencia) tiene 4 rutas. Sus horas de operación los días laborables regulares son: (1) a 265a Valencia: 7:00 - 17:40 (2) a 265a Yatova (Directo): 9:00 - 19:45 (3) a 265b Yatova (Por Cheste): 7:30 - 21:00 (4) a Valencia (Por Cheste): 5:50 - 19:10 Usa la aplicación Moovit para encontrar la parada de la línea 265 de autobús más cercana y descubre cuándo llega la próxima línea 265 de autobús Sentido: 265a Valencia Horario de la línea 265 de autobús 25 paradas 265a Valencia Horario de ruta: VER HORARIO DE LA LÍNEA lunes 7:00 - 17:40 martes 7:00 - 17:40 Paseo De La Alberca 5 Cl Grao, Yátova miércoles 7:00 - 17:40 Dr. Peset Alexandre 7 jueves 7:00 - 17:40 2 Cl Dr Peset Aleixandre, Yátova viernes 7:00 - 17:40 Avenida Del Pais Valenciano 1 sábado Sin servicio 27 Av Hoya De Buñol, Macastre domingo Sin servicio Plaza De España 6 Pz España, Alborache Valencia 28 2 Cl Grao, Alborache Información de la línea 265 de autobús Dirección: 265a Valencia Beltrán Báguena Paradas: 25 3 Av Beltran Baguena, Buñol Duración del viaje: 75 min Resumen de la línea: Paseo De La Alberca, Dr. Peset Jaime Balmes 6 [Buñol] Alexandre 7, Avenida Del Pais Valenciano 1, Plaza De 16 Cl Balmes, Buñol España, Valencia 28, Beltrán Báguena, Jaime Balmes 6 [Buñol], Blasco Ibáñez-24, Av Blasco Blasco Ibáñez-24 Ibáñez (Supermercat) [Buñol], Polígon Pailla [Chiva], 24 Av Blasco Ibañez, Buñol Ramón Y Cajal - La Báscula, Plaça Constitucion 3 [Chiva], Maestro García Navarro 26, A - 3 (Vía De Av Blasco Ibáñez (Supermercat) [Buñol] Servicio), A - 3 (Vía De Servicio), Venta Del Poyo 70 Av Blasco Ibañez, Buñol [Riba-Roja De Túria], Arquitecto Salvador Pascual, Comarques Pais Valencia - La Cautiva [Quart De Polígon Pailla [Chiva] Poblet], Cid - Barri De La Llum, Cid (Illeta) - Tres 93 Pl Pahilla, Chiva Creus, Cid- Parque Oeste [València], San Francisco De Borja, Plaza España, G.V.