Plant Propagation Protocol for Corallorhiza Striata Lindl., Orth. Var
Total Page:16
File Type:pdf, Size:1020Kb
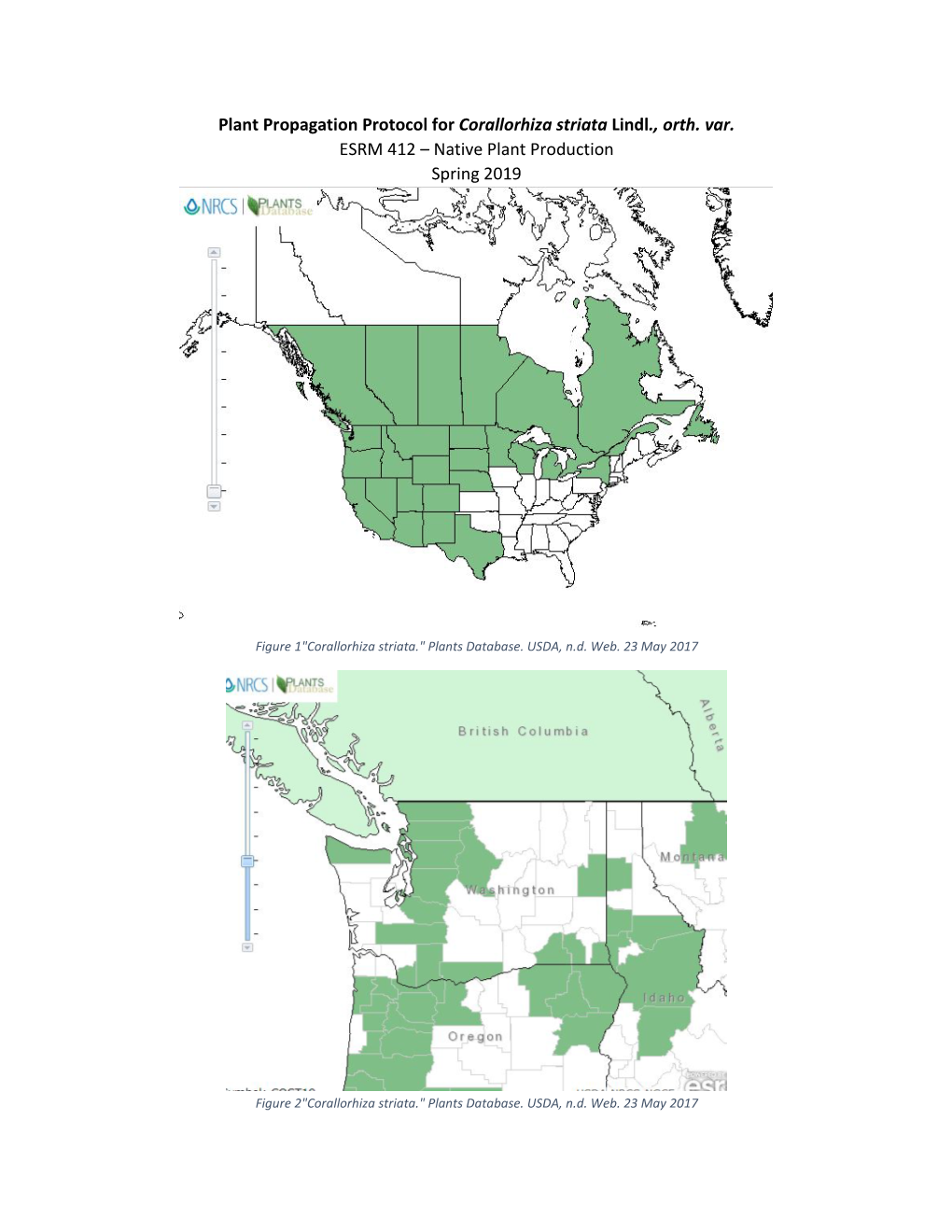
Load more
Recommended publications
-

Orchids of Lakeland
a field guide to OrchidsLakeland Provincial Park, Provincial Recreation Area and surrounding region Written by: Patsy Cotterill Illustrated by: John Maywood Pub No: I/681 ISBN: 0-7785-0020-9 Printed 1998 Sepal Sepal Petal Petal Seed Sepal Lip (labellum) Ovary Finding an elusive (capsule) orchid nestled away in Sepal the boreal forest is a Scape thrilling experience for amateur and Petal experienced naturalists alike. This type Petal of recreation has become a passion for Column some and, for many others, a wonder- ful way to explore nature and learn Ovary Sepal (capsule) more about the living world around us. Sepal But, we also need to be careful when Lip (labellum) looking at these delicate plants. Be mindful of their fragile nature... keep Spur only memories and take only photo- graphs so that others, too, can enjoy. Corm OrchidsMany people are surprisedin Alberta to learn that orchids grow naturally in Alberta. Orchids are typically associated with tropical rain forests (where indeed, many do grow) or flower-shop purchases for special occasions, exotic, expensive and highly ornamental. Wild orchids in fact are found world-wide (except in Antarctica). They constitute Roots Tuber the second largest family of flowering plants (the Orchidaceae) after the Aster family, with upwards of 15,000 species. In Alberta, there are 26 native species. Like all Roots orchids from north temperate regions, these are terrestrial. They grow in the ground, in contrast to the majority of rain forest species which are epiphytes, that is, they grow attached to trees for support but do not derive nourishment from them. -

Plant Genetics
RESEARCH HIGHLIGHTS Nature Reviews Genetics | AOP, published online 9 October 2014; doi:10.1038/nrg3843 PLANT GENETICS Notably, in accordance with the team’s earlier hypothesis on the specific stepwise progression of gene loss in Following the early root heterotrophic flowering plants (NAD(P)H genes followed by photosynthesis-related genes, of plastome degradation plastid-encoded RNA polymerase genes, ATP synthase genes and Parasitic organisms are often for high-throughput sequencing. housekeeping genes), coralroots genomically and morphologically De novo assembly was then used to are in the early stages of genome reduced. For example, some parasitic piece together the genomes and degradation. This study indicates that coralroots plants contain degraded plastid to define genes and pseudogenes. a specific path does seem to exist for are in the genomes owing to a reduced need In addition, chlorophyll content was plastid genome degradation (with to photosynthesize. A new study measured for several plants from some exceptions) and that coralroots initial steps now shows that coralroot orchids are each species. Some coralroot orchids are in the initial steps towards towards the in the early stages of the transition have green tissues, and it is thought the transition to a highly reduced transition to a to a parasitic lifestyle, and that this that these species are still able to chloroplast genome, which is typical condition has arisen independently at photosynthesize, whereas those of obligate parasites. “This condition highly reduced least twice in the genus. without any green tissues cannot. has arisen independently at least chloroplast Most coralroot orchids do not have The authors report a correlation twice in the genus, making coralroots genome leaves, produce little chlorophyll between the number of functional a powerful system in which to study and depend on fungi to obtain chloroplast genes and chlorophyll the transition to parasitism,” explains nutrients. -

The Vascular Flora of Rarău Massif (Eastern Carpathians, Romania). Note Ii
Memoirs of the Scientific Sections of the Romanian Academy Tome XXXVI, 2013 BIOLOGY THE VASCULAR FLORA OF RARĂU MASSIF (EASTERN CARPATHIANS, ROMANIA). NOTE II ADRIAN OPREA1 and CULIŢĂ SÎRBU2 1 “Anastasie Fătu” Botanical Garden, Str. Dumbrava Roşie, nr. 7-9, 700522–Iaşi, Romania 2 University of Agricultural Sciences and Veterinary Medicine Iaşi, Faculty of Agriculture, Str. Mihail Sadoveanu, nr. 3, 700490–Iaşi, Romania Corresponding author: [email protected] This second part of the paper about the vascular flora of Rarău Massif listed approximately half of the whole number of the species registered by the authors in their field trips or already included in literature on the same area. Other taxa have been added to the initial list of plants, so that, the total number of taxa registered by the authors in Rarău Massif amount to 1443 taxa (1133 species and 310 subspecies, varieties and forms). There was signaled out the alien taxa on the surveyed area (18 species) and those dubious presence of some taxa for the same area (17 species). Also, there were listed all the vascular plants, protected by various laws or regulations, both internal or international, existing in Rarău (i.e. 189 taxa). Finally, there has been assessed the degree of wild flora conservation, using several indicators introduced in literature by Nowak, as they are: conservation indicator (C), threat conservation indicator) (CK), sozophytisation indicator (W), and conservation effectiveness indicator (E). Key words: Vascular flora, Rarău Massif, Romania, conservation indicators. 1. INTRODUCTION A comprehensive analysis of Rarău flora, in terms of plant diversity, taxonomic structure, biological, ecological and phytogeographic characteristics, as well as in terms of the richness in endemics, relict or threatened plant species was published in our previous note (see Oprea & Sîrbu 2012). -

Phylogeny, Character Evolution and the Systematics of Psilochilus (Triphoreae)
THE PRIMITIVE EPIDENDROIDEAE (ORCHIDACEAE): PHYLOGENY, CHARACTER EVOLUTION AND THE SYSTEMATICS OF PSILOCHILUS (TRIPHOREAE) A Dissertation Presented in Partial Fulfillment of the Requirements for The Degree Doctor of Philosophy in the Graduate School of the Ohio State University By Erik Paul Rothacker, M.Sc. ***** The Ohio State University 2007 Doctoral Dissertation Committee: Approved by Dr. John V. Freudenstein, Adviser Dr. John Wenzel ________________________________ Dr. Andrea Wolfe Adviser Evolution, Ecology and Organismal Biology Graduate Program COPYRIGHT ERIK PAUL ROTHACKER 2007 ABSTRACT Considering the significance of the basal Epidendroideae in understanding patterns of morphological evolution within the subfamily, it is surprising that no fully resolved hypothesis of historical relationships has been presented for these orchids. This is the first study to improve both taxon and character sampling. The phylogenetic study of the basal Epidendroideae consisted of two components, molecular and morphological. A molecular phylogeny using three loci representing each of the plant genomes including gap characters is presented for the basal Epidendroideae. Here we find Neottieae sister to Palmorchis at the base of the Epidendroideae, followed by Triphoreae. Tropidieae and Sobralieae form a clade, however the relationship between these, Nervilieae and the advanced Epidendroids has not been resolved. A morphological matrix of 40 taxa and 30 characters was constructed and a phylogenetic analysis was performed. The results support many of the traditional views of tribal composition, but do not fully resolve relationships among many of the tribes. A robust hypothesis of relationships is presented based on the results of a total evidence analysis using three molecular loci, gap characters and morphology. Palmorchis is placed at the base of the tree, sister to Neottieae, followed successively by Triphoreae sister to Epipogium, then Sobralieae. -

Second Contribution to the Vascular Flora of the Sevastopol Area
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Wulfenia Jahr/Year: 2015 Band/Volume: 22 Autor(en)/Author(s): Seregin Alexey P., Yevseyenkow Pavel E., Svirin Sergey A., Fateryga Alexander Artikel/Article: Second contribution to the vascular flora of the Sevastopol area (the Crimea) 33-82 © Landesmuseum für Kärnten; download www.landesmuseum.ktn.gv.at/wulfenia; www.zobodat.at Wulfenia 22 (2015): 33 – 82 Mitteilungen des Kärntner Botanikzentrums Klagenfurt Second contribution to the vascular flora of the Sevastopol area (the Crimea) Alexey P. Seregin, Pavel E. Yevseyenkov, Sergey A. Svirin & Alexander V. Fateryga Summary: We report 323 new vascular plant species for the Sevastopol area, an administrative unit in the south-western Crimea. Records of 204 species are confirmed by herbarium specimens, 60 species have been reported recently in literature and 59 species have been either photographed or recorded in field in 2008 –2014. Seventeen species and nothospecies are new records for the Crimea: Bupleurum veronense, Lemna turionifera, Typha austro-orientalis, Tyrimnus leucographus, × Agrotrigia hajastanica, Arctium × ambiguum, A. × mixtum, Potamogeton × angustifolius, P. × salicifolius (natives and archaeophytes); Bupleurum baldense, Campsis radicans, Clematis orientalis, Corispermum hyssopifolium, Halimodendron halodendron, Sagina apetala, Solidago gigantea, Ulmus pumila (aliens). Recently discovered Calystegia soldanella which was considered to be extinct in the Crimea is the most important confirmation of historical records. The Sevastopol area is one of the most floristically diverse areas of Eastern Europe with 1859 currently known species. Keywords: Crimea, checklist, local flora, taxonomy, new records A checklist of vascular plants recorded in the Sevastopol area was published seven years ago (Seregin 2008). -

Striped Coralroot (Corallorhiza Striata) Did You Know?
Striped Coralroot (Corallorhiza striata) Did you Know? The striped Coralroot is the largest of all the Coralroots and is shared with the family of orchids. 44 of the 65 orchids in Canada call Ontario home, and can be seen on the Escarpment. This plant has no leaves. It is classified as a saprophytic plant, meaning it has no chlorophyll and relies on fungi for nutrients and energy. Habitat – Flourishes in coniferous and deciduous forests, especially open woods and very shade tolerant. Size – Can grow up to 60 cm (24 in) high, with flowers of about 1.5 cm in size. Range – Canada (excluding northern provinces), and can be found in almost all of the United States. Status – Ontario Status: S4,Secure. Canadian Status: Secure. The Bruce Trail Conservancy | PO Box 857 Hamilton, ON L8N 3N9 | 1.800.665.4453 | [email protected] Identification: These orchids are the largest of the coralroots. The striped coralroot can be noted by the pale pinkish- white sepals and petals, or ruby red lips that grow along its red herbaceous stem with maroon thin stripes. There are red or purple spots inside each flower, marking the male and female parts. Typically it grows in clumps, rarely exceeding 40 cm in height in Ontario. Interesting Facts This plant does not contain chlorophyll, therefore photosynthesis cannot occur. Instead, the Striped Coralroot is dependent on getting its energy and nutrients from mycorrhizal fungi. The fungus infects its roots and gathers nutrients from surrounding organic matter. This is also the reason why this plant is not green. This species got its name because the roots resemble coral, and Corallorhiza translates into “coral root” in Greek, and Striata meaning “striped”. -

Pterospora Andromedea Nutt
Pterospora andromedea Nutt. pine-drops State Distribution Best Survey Period Photo by Daniel C. Nepstad Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Status: State threatened indicated as ‘rare.’ Seventeen occurrences occur on public lands or designated preserves. None of these occurrences, Global and state rank: G5/S2 however, are under specific active protection. Family: Monotropaceae (Indian-pipe) Recognition: Pine-drop lacks chlorophyll and has one to several simple, erect stems, from 3-10 dm tall, bearing Other common names: giant birds nest, Albany beech- numerous scale-like leaves and a terminal raceme of drops numerous nodding flowers. The approx. 6-7 mm long , Synonyms: Monotropa procera Torr. bell-shaped corolla is white while the sepals and vegeta- tive parts of the plant are reddish to maroon. The stem Taxonomy: Pterospora andromeda is the only species in and sepals are glandular-hairy giving the plant a its genus (monotypic). Sometimes included in the clammy-sticky feel. The similar, but more widespread and Pyrolaceae or Ericaceae under subfamily Pyrolaceae, common species Monotropa uniflora (Indian pipe) and M. Pterospora and other species of the Monotropaceae differ hypopithys (pinesap), also lack chlorophyll, but are in their saprophytic (absorb nutrients from dead or decay- typically one half the size of Pterospora or smaller. In ing matter) habit (Voss 1996). addition, the flowers of both Indian pipe and pinesap become erect in fruit, unlike the strongly nodding fruits of Total range: A species primarily of Western North Pterospora. Indian pipe also differs in bearing only a America, pine drops is disjunct in the Great Lakes region single large flower on each stem. -

Ecosystem-Based Management Plan for Cypress Hills Interprovincial Park
Cypress Hills Interprovincial Park ECOSYSTEM-BASED MANAGEMENT PLAN Saskatchewan.ca Ecosystem-Based Management Plan February 19, 2020 Cypress Hills Interprovincial Park ECOSYSTEM-BASED MANAGEMENT PLAN PROJECT REFERENCE NUMBER: 1467-5 March 11, 2020 Revised: March 2021 Prepared for: Saskatchewan Ministry of Parks, Culture and Sport 3211 Albert St Regina, SK S4S 5W6 Cypress Hills Interprovincial Park Page | 1 Approval Form The Ecosystem-based Management Plan for Cypress Hills Interprovincial Park (2020) is hereby approved for use by the Ministry of Parks, Culture and Sport in the management of the ecosystem and landscape of Cypress Hills Interprovincial Park. March 11, 2020 Darryl Sande, RPF, Plan Author Date FORSITE Inc. Recommended for approval by: March 1, 2021 Thuan Chu, Senior Park Landscape Ecologist Date Landscape Protection Unit Ministry of Parks, Culture and Sport Cypress Hills Interprovincial Park Page | ii EXECUTIVE SUMMARY Cypress Hills Interprovincial Park (CHIPP) is a 183 square kilometre Natural Environment Park within the southwestern corner of Saskatchewan. The park encompasses the unique geological features and elevation of the Cypress Hills formation. The area contains a mix of Boreal and Montane forest elements as well as Prairie grassland elements. The area is surrounded by agricultural and pasture lands. The park is made up of a mix of natural forests and grasslands, which is classified into nine ecosites. Upland ecosites include plains rough fescue grassland on silty clay loam, lodgepole pine-dominated stands on sandy clay, white spruce stands on silty clay, aspen stands on clay loam, aspen-white spruce mixedwoods on silty clay soils, and aspen-lodgepole pine mixedwoods on clay loam. -

Systematics, Phylogeography, Fungal Associations, and Photosynthesis
Systematics, Phylogeography, Fungal Associations, and Photosynthesis Gene Evolution in the Fully Mycoheterotrophic Corallorhiza striata Species Complex (Orchidaceae: Epidendroideae) Dissertation Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of the Ohio State University By Craig Francis Barrett, M. S. Evolution, Ecology, and Organismal Biology ***** The Ohio State University 2010 Dissertation Committee: Dr. John V. Freudenstein, Advisor Dr. John W. Wenzel Dr. Andrea D. Wolfe Copyright by Craig Francis Barrett 2010 ABSTRACT Corallorhiza is a genus of obligately mycoheterotrophic (fungus-eating) orchids that presents a unique opportunity to study phylogeography, taxonomy, fungal host specificity, and photosynthesis gene evolution. The photosysnthesis gene rbcL was sequenced for nearly all members of the genus Corallorhiza; evidence for pseudogene formation was found in both the C. striata and C. maculata complexes, suggesting multiple independent transitions to complete heterotrophy. Corallorhiza may serve as an exemplary system in which to study the plastid genomic consequences of full mycoheterotrophy due to relaxed selection on photosynthetic apparatus. Corallorhiza striata is a highly variable species complex distributed from Mexico to Canada. In an investigation of molecular and morphological variation, four plastid DNA clades were identified, displaying statistically significant differences in floral morphology. The biogeography of C. striata is more complex than previously hypothesized, with two main plastid lineages present in both Mexico and northern North America. These findings add to a growing body of phylogeographic data on organisms sharing this common distribution. To investigate fungal host specificity in the C. striata complex, I sequenced plastid DNA for orchids and nuclear DNA for fungi (n=107 individuals), and found that ii the four plastid clades associate with divergent sets of ectomycorrhizal fungi; all within a single, variable species, Tomentella fuscocinerea. -

NORTH AMERICAN NATIVE ORCHID JOURNAL Volume 15(1) 2009
NORTH AMERICAN NATIVE ORCHID JOURNAL Volume 15(1) 2009 IN THIS ISSUE: A FAMILY ORCHID VACATION TO THE GREAT LAKES REGION AND POINTS BEYOND MORPHOLOGICAL VARIATION IN HABENARIA MACROCERATITIS PLATANTHERA HYBRIDS FROM WESTERN NORTH AMERICA TWO NEW FORMS OF THE FLORIDA ADDER’S-MOUTH A NEW GENUS FOR THE NORTH AMERICAN CLEISTES NEW COMBINATIONS AND A NEW SPECIES IN CLEISTESIOPSIS and more…………. The North American Native Orchid Journal (ISSN 1084-7332) is a publication devoted to promoting interest and knowledge of the native orchids of North America. A limited number of the print version of each issue of the Journal are available upon request and electronic versions are available to all interested persons or institutions free of charge. The Journal welcomes articles of any nature that deal with native or introduced orchids that are found growing wild in North America, primarily north of Mexico, although articles of general interest concerning Mexican species will always be welcome. NORTH AMERICAN NATIVE ORCHID JOURNAL Volume 15 (1) 2009 CONTENTS NOTES FROM THE EDITOR 1 A FAMILY ORCHID VACATION TO THE GREAT LAKES REGION AND POINTS BEYOND TOM NELSON 2 MORPHOLOGICAL VARIATION IN HABENARIA MACROCERATITIS SCOTT L. STEWART, PHD. 36 NEW TAXA& COMBINATIONS P.M. BROWN FOUR NEW PLATANTHERA HYBRIDS FROM WESTERN NORTH AMERICA 40 TWO NEW FORMS OF THE FLORIDA ADDER’S-MOUTH, MALAXIS SPICATA 43 A WHITE/GREEN FORM OF THE GENTIAN NODDINGCAPS, TRIPHORA GENTIANOIDES 45 WHAT I DID ON MY SUMMER VACATION THE SLOW EMPIRICIST 46 A NEW GENUS FOR THE NORTH AMERICAN CLEISTES E. PANSARIN ET AL. 50 NEW COMBINATIONS AND A NEW SPECIES IN CLEISTESIOPSIS P.M. -

Diversity and Evolution of Monocots
Lilioids - petaloid monocots 4 main groups: Diversity and Evolution • Acorales - sister to all monocots • Alismatids of Monocots – inc. Aroids - jack in the pulpit ! • Lilioids (lilies, orchids, yams) – grade, non-monophyletic – petaloid . orchids and palms . ! • Commelinoids – Arecales – palms – Commelinales – spiderwort – Zingiberales –banana – Poales – pineapple – grasses & sedges Lilioids - petaloid monocots Asparagales: *Orchidaceae - orchids • finish the Asparagales by 1. Terrestrial/epiphytes: plants looking at the largest family - typically not aquatic the orchids 2. Geophytes: herbaceous above ground with below ground modified perennial stems: bulbs, corms, rhizomes, tubers 3. Tepals: showy perianth in 2 series of 3 each; usually all petaloid, or outer series not green and sepal-like & with no bracts 1 *Orchidaceae - orchids *Orchidaceae - orchids The family is diverse with about 880 genera and over 22,000 All orchids have a protocorm - a feature restricted to the species, mainly of the tropics family. Orchids are • structure formed after germination and before the mycotrophic (= fungi development of the seedling plant dependent) lilioids; • has no radicle but instead mycotrophic tissue some are obligate mycotrophs Cypripedium acaule Corallorhiza striata Stemless lady-slipper Striped coral root Dactylorhiza majalis protocorm *Orchidaceae - orchids *Orchidaceae - orchids Cosmopolitan, but the majority of species are found in the Survive in these epiphytic and other harsh environments via tropics and subtropics, ranging from sea -

Liliales) Constantijn B
Journal of Biogeography (J. Biogeogr.) (2015) ORIGINAL Ancient Gondwana break-up explains the ARTICLE distribution of the mycoheterotrophic family Corsiaceae (Liliales) Constantijn B. Mennes1,*, Vivienne K. Y. Lam2, Paula J. Rudall3, Stephanie P. Lyon4, Sean W. Graham2, Erik F. Smets1,5 and Vincent S. F. T. Merckx1 1Naturalis Biodiversity Center, Leiden ABSTRACT University, Leiden, The Netherlands, Aim Many plant families have a disjunct distribution across the southern Paci- 2Department of Botany, University of British fic Ocean, including the mycoheterotrophic family Corsiaceae, which provides Columbia, Vancouver, British Columbia V6T 1Z4, Canada, 3Royal Botanic Gardens Kew, a prime example of this biogeographical pattern. A better grasp of the family’s Richmond, Surrey, UK, 4Department of evolutionary relationships is needed to understand its historical biogeography. Botany, University of Wisconsin Madison, We therefore aimed to (1) test the uncertain monophyly of Corsiaceae, (2) Madison, WI 54706, USA, 5Section Ecology, define its phylogenetic position, and (3) estimate divergence times for the fam- Evolution and Biodiversity Conservation, KU ily, allowing us to assess whether the distribution of the family is the result of Leuven, BE-3001 Leuven, Belgium vicariance. Location Southern South America and Australasia. Methods We analysed various combinations of mitochondrial and nuclear data to address the monophyly, phylogenetic position and age of Corsiaceae. To test its monophyly, we used a three-locus data set including most monocot orders, and to infer its exact phylogenetic position, we used a five-locus extended data set. We corroborated these findings using an independent plas- tome dataset. We then used a two-locus dataset with taxa from all monocot orders, and a three-locus dataset containing only taxa of Liliales, to estimate divergence times using a fossil-calibrated uncorrelated lognormal relaxed-clock approach.