Cortical Area V4 and Its Role in the Perception of Color
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
Chapter 6 COLOR and COLOR VISION
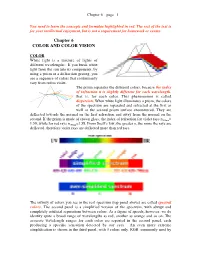
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

Rethinking Color Cameras
Rethinking Color Cameras Ayan Chakrabarti William T. Freeman Todd Zickler Harvard University Massachusetts Institute of Technology Harvard University Cambridge, MA Cambridge, MA Cambridge, MA [email protected] [email protected] [email protected] Abstract Digital color cameras make sub-sampled measurements of color at alternating pixel locations, and then “demo- saick” these measurements to create full color images by up-sampling. This allows traditional cameras with re- stricted processing hardware to produce color images from a single shot, but it requires blocking a majority of the in- cident light and is prone to aliasing artifacts. In this paper, we introduce a computational approach to color photogra- phy, where the sampling pattern and reconstruction process are co-designed to enhance sharpness and photographic speed. The pattern is made predominantly panchromatic, thus avoiding excessive loss of light and aliasing of high spatial-frequency intensity variations. Color is sampled at a very sparse set of locations and then propagated through- out the image with guidance from the un-aliased luminance channel. Experimental results show that this approach often leads to significant reductions in noise and aliasing arti- facts, especially in low-light conditions. Figure 1. A computational color camera. Top: Most digital color cameras use the Bayer pattern (or something like it) to sub-sample 1. Introduction color alternatingly; and then they demosaick these samples to create full-color images by up-sampling. Bottom: We propose The standard practice for one-shot digital color photog- an alternative that samples color very sparsely and is otherwise raphy is to include a color filter array in front of the sensor panchromatic. -

Spatial Filtering, Color Constancy, and the Color-Changing Dress Department of Psychology, American University, Erica L
Journal of Vision (2017) 17(3):7, 1–20 1 Spatial filtering, color constancy, and the color-changing dress Department of Psychology, American University, Erica L. Dixon Washington, DC, USA Department of Psychology and Department of Computer Arthur G. Shapiro Science, American University, Washington, DC, USA The color-changing dress is a 2015 Internet phenomenon divide among responders (as well as disagreement from in which the colors in a picture of a dress are reported as widely followed celebrity commentators) fueled a rapid blue-black by some observers and white-gold by others. spread of the photo across many online news outlets, The standard explanation is that observers make and the topic trended worldwide on Twitter under the different inferences about the lighting (is the dress in hashtag #theDress. The huge debate on the Internet shadow or bright yellow light?); based on these also sparked debate in the vision science community inferences, observers make a best guess about the about the implications of the stimulus with regard to reflectance of the dress. The assumption underlying this individual differences in color perception, which in turn explanation is that reflectance is the key to color led to a special issue of the Journal of Vision, for which constancy because reflectance alone remains invariant this article is written. under changes in lighting conditions. Here, we The predominant explanations in both scientific demonstrate an alternative type of invariance across journals (Gegenfurtner, Bloj, & Toscani, 2015; Lafer- illumination conditions: An object that appears to vary in Sousa, Hermann, & Conway, 2015; Winkler, Spill- color under blue, white, or yellow illumination does not change color in the high spatial frequency region. -

Color Constancy and Contextual Effects on Color Appearance
Chapter 6 Color Constancy and Contextual Effects on Color Appearance Maria Olkkonen and Vebjørn Ekroll Abstract Color is a useful cue to object properties such as object identity and state (e.g., edibility), and color information supports important communicative functions. Although the perceived color of objects is related to their physical surface properties, this relationship is not straightforward. The ambiguity in perceived color arises because the light entering the eyes contains information about both surface reflectance and prevailing illumination. The challenge of color constancy is to estimate surface reflectance from this mixed signal. In addition to illumination, the spatial context of an object may also affect its color appearance. In this chapter, we discuss how viewing context affects color percepts. We highlight some important results from previous research, and move on to discuss what could help us make further progress in the field. Some promising avenues for future research include using individual differences to help in theory development, and integrating more naturalistic scenes and tasks along with model comparison into color constancy and color appearance research. Keywords Color perception • Color constancy • Color appearance • Context • Psychophysics • Individual differences 6.1 Introduction Color is a useful cue to object properties such as object identity and state (e.g., edibility), and color information supports important communicative functions [1]. Although the perceived color of objects is related to their physical surface properties, M. Olkkonen, M.A. (Psych), Dr. rer. nat. (*) Department of Psychology, Science Laboratories, Durham University, South Road, Durham DH1 3LE, UK Institute of Behavioural Sciences, University of Helsinki, Siltavuorenpenger 1A, 00014 Helsinki, Finland e-mail: [email protected]; maria.olkkonen@helsinki.fi V. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

1 Human Color Vision
CAMC01 9/30/04 3:13 PM Page 1 1 Human Color Vision Color appearance models aim to extend basic colorimetry to the level of speci- fying the perceived color of stimuli in a wide variety of viewing conditions. To fully appreciate the formulation, implementation, and application of color appearance models, several fundamental topics in color science must first be understood. These are the topics of the first few chapters of this book. Since color appearance represents several of the dimensions of our visual experience, any system designed to predict correlates to these experiences must be based, to some degree, on the form and function of the human visual system. All of the color appearance models described in this book are derived with human visual function in mind. It becomes much simpler to understand the formulations of the various models if the basic anatomy, physiology, and performance of the visual system is understood. Thus, this book begins with a treatment of the human visual system. As necessitated by the limited scope available in a single chapter, this treatment of the visual system is an overview of the topics most important for an appreciation of color appearance modeling. The field of vision science is immense and fascinating. Readers are encouraged to explore the liter- ature and the many useful texts on human vision in order to gain further insight and details. Of particular note are the review paper on the mechan- isms of color vision by Lennie and D’Zmura (1988), the text on human color vision by Kaiser and Boynton (1996), the more general text on the founda- tions of vision by Wandell (1995), the comprehensive treatment by Palmer (1999), and edited collections on color vision by Backhaus et al. -

Color Vision Deficiency
Color Vision Deficiency What is color vision deficiency? Color vision deficiency is called “color blindness” by mistake. Actually, the term describes a number of different problems people have with color vision. Abnormal color vision may vary from not being able to tell certain colors apart to not being able to identify any color. Whom does color vision deficiency affect? An estimated 8% of males and fewer than 1% of females have color vision problems. Most color vision problems run in families and are inherited and present at birth. A child inherits a color vision deficiency by receiving a faulty color vision gene from a parent. Abnormal color vision is found in a recessive gene on the X chromosome. Men are born with just one X and one Y chromosome. However, women have two X chromosomes. Because of this, women can sometimes overcome the faulty gene with their second normal X chromosome. Men, unfortunately, do not have a second X chromosome to help compensate for the faulty color vision gene. Heredity does not cause all color vision problems. One common problem happens from the normal aging of the eye’s lens. The lens is clear at birth, but the aging process causes it to darken and yellow. Older adults may have problems identifying certain dark colors, particularly blues. Certain medications as well as inherited or acquired retinal and optic nerve disease, may also affect normal color vision. Who should be tested for color deficiency? Any child who is having difficulty in school should be checked for possible visual problems including color vision impairment. -

Fast Fourier Color Constancy
Fast Fourier Color Constancy Jonathan T. Barron Yun-Ta Tsai [email protected] [email protected] Abstract based techniques in computer vision, both problems reduce to just estimating the “best” illuminant from an image, and We present Fast Fourier Color Constancy (FFCC), a the question of whether that illuminant is objectively true color constancy algorithm which solves illuminant esti- or subjectively attractive is just a matter of the data used mation by reducing it to a spatial localization task on a during training. torus. By operating in the frequency domain, FFCC pro- Despite their accuracy, modern learning-based color duces lower error rates than the previous state-of-the-art by constancy algorithms are not immediately suitable as prac- 13 − 20% while being 250 − 3000× faster. This unconven- tical white balance algorithms, as practical white balance tional approach introduces challenges regarding aliasing, has several requirements besides accuracy: directional statistics, and preconditioning, which we ad- Speed - An algorithm running in a camera’s viewfinder dress. By producing a complete posterior distribution over must run at 30 FPS on mobile hardware. But a camera’s illuminants instead of a single illuminant estimate, FFCC compute budget is precious: demosaicing, face detection, enables better training techniques, an effective temporal auto exposure, etc, must also run simultaneously and in real smoothing technique, and richer methods for error analy- time. Spending more than a small fraction (say, 5 − 10%) sis. Our implementation of FFCC runs at ∼ 700 frames per of a camera’s compute budget on white balance is impracti- second on a mobile device, allowing it to be used as an ac- cal, suggesting that our speed requirement is closer to 1 − 5 curate, real-time, temporally-coherent automatic white bal- milliseconds per frame. -

Color Vision Mechanisms
11 COLOR VISION MECHANISMS Andrew Stockman Department of Visual Neuroscience UCL Institute of Opthalmology London, United KIngdom David H. Brainard Department of Psychology University of Pennsylvania Philadelphia, Pennsylvania 11.1 GLOSSARY Achromatic mechanism. Hypothetical psychophysical mechanisms, sometimes equated with the luminance mechanism, which respond primarily to changes in intensity. Note that achromatic mech- anisms may have spectrally opponent inputs, in addition to their primary nonopponent inputs. Bezold-Brücke hue shift. The shift in the hue of a stimulus toward either the yellow or blue invariant hues with increasing intensity. Bipolar mechanism. A mechanism, the response of which has two mutually exclusive types of out- put that depend on the balance between its two opposing inputs. Its response is nulled when its two inputs are balanced. Brightness. A perceptual measure of the apparent intensity of lights. Distinct from luminance in the sense that lights that appear equally bright are not necessarily of equal luminance. Cardinal directions. Stimulus directions in a three-dimensional color space that silence two of the three “cardinal mechanisms.” These are the isolating directions for the L+M, L–M, and S–(L+M) mech- anisms. Note that the isolating directions do not necessarily correspond to mechanism directions. Cardinal mechanisms. The second-site bipolar L–M and S–(L+M) chromatic mechanisms and the L+M luminance mechanism. Chromatic discrimination. Discrimination of a chromatic target from another target or back- ground, typically measured at equiluminance. Chromatic mechanism. Hypothetical psychophysical mechanisms that respond to chromatic stimuli, that is, to stimuli modulated at equiluminance. Color appearance. Subjective appearance of the hue, brightness, and saturation of objects or lights. -

38 Color Constancy
38 Color Constancy DAVID H. BRAINARD AND ANA RADONJIC Vision is useful because it informs us about the physical provide a self-contained introduction, to highlight environment. In the case of colm~ two distinct functions some more recent results, and to outline what we see are generally emphasized (e.g., Jacobs, 1981; Mollon, as important challenges for the field. We focus on key 1989). First, color helps to segment objects from each concepts and avoid much in the way of technical devel other and the background. A canonical task for this opment. Other recent treatments complement the one function is locating fruit in foliage (Regan et al., 2001; provided here and provide a level of technical detail Sumner & Mollon, 2000). Second, color provides infm' beyond that introduced in this chapter (Brainard, 2009; mation about object properties (e.g., fresh fish versus Brainard & Maloney, 2011; Foster, 2011; Gilchrist, 2006; old fish; figure 38.1). Using color for this second func Kingdom, 2008; Shevell & Kingdom, 2008; Smithson, tion is enabled to the extent that perceived object color 2005; Stockman & Brainard, 2010). correlates well with object reflectance properties. Achieving such correlation is a nontrivial requirement, MEASURING CONSTANCY however, because the spectrum of the light reflected from an object confounds variation in object surface The study of constancy requires methods for measuring reflectance with variation in the illumination (figure it. The classic approach to such measurement is to 38.2; Brainard, Wandell, & Chichilnisky, 1993; Hurl assess the extent to which the color appearance of bert, 1998; Maloney, 1999). In particular, the spectrum objects varies as they are viewed under different illumi of the reflected light is the wavelength-by-wavelength nations. -

Color Blindness Fovea
Color Theory • Chapter 3b Perceiving Color • Color Constancy • Chapter 3 Perceiving Color Theory Color (so far) • Lens • Iris • Rods • Cones x3 • Fovea • Optic Nerve • Iodopsin & Rhodopsin • Chapter 3 Color Theory Perceiving Color (today) • Tri-Chromatic theory of color vision • Color Afterimage • Opponent Theory of color vision • Color Constancy • Monet’s series • Color Blindness Fovea • A tiny (1mm) area on the retina is covered with a dense collection of cones. • This region is where we see color distinctions best. Seeing Color • Still no proven model of how color perception works. • Cones contain light-sensitive pigments called Iodopsin. • Iodopsin changes when light hits it -- it is photo- sensitive. Light is effectively converted/translated into a nerve impulse…a signal to the brain. Trichromatic Theory & Opponent Theory • 19th c. Trichromatic Theory first proposed the idea of three types of cones. • Current theory -- Opponent Theory -- is that there are three types of iodopsin -- one that senses (or “sees”) red light, one that sees green, and one blue-violet. Each is “blind” to its complement. • We then combine the information from all three to perceive color. • We have mostly red- and green- sensitive cones – few blue-sensitive ones. • Diagram of suspected “wiring” of cones to ganglion Cones to Color (nerve) cells. • Light primaries are “read” individually, then results are combined. Cones to Color • Diagram of suspected trichromatic “wiring” of cones to ganglion (nerve) cells. • Light primaries are “read” individually, then results are combined. • http://www.handprint.com/HP /WCL/color1.html Opponent Processing • “As the diagram shows, the opponent processing pits the responses from one type of cone against the others. -

Color Vision
Color Vision Lecture 10 Chapter 5, Part A Jonathan Pillow Sensation & Perception (PSY 345 / NEU 325) Princeton University, Fall 2017 1 Exam #1: Thursday 10/19 Format: multiple-choice, fill-in-the-blank, & short answer What to study: - all material from lectures & slides - precept readings (basic gist & findings of each article) (If something appeared only in the book, and not at all in class or precept or slides, you can probably safely ignore it) Review session: tonight @ 7:00 in PNI A30 2 2015 called…. Grey-and-green, or pink-and-white? 3 4 5 • color vision has evolutionary value • lack of color vision ≠ black & white 6 Basic Principles of Color Perception The book says: “Color is not a physical property but a psychophysical property” 7 Basic Principles of Color Perception • Most of the light we see is reflected • Typical light sources: Sun, light bulb, LED screen • We see only part of the electromagnetic spectrum(between 400 and 700 nm). Why?? 8 Basic Principles of Color Perception • Why only 400-700 nm? (Pomerantz, Rice U.) Suggestion: unique ability to penetrate sea water 9 Basic Principles of Color Perception Q: How many numbers would you need to write down to specify the spectral properties of a light source? A: It depends on how you “bin” up the spectrum • One number for each spectral “bin”: 20 17 16 15 13 12 10 example: 13 5 bins energy 0 0 0 0 0 10 Basic Principles of Color Perception Device: hyper-spectral camera - measures the amount of energy (or number of photons) in each small range of wavelengths - can use thousands of bins (or “frequency bands”) instead of just the 13 shown here 20 17 16 15 13 12 10 5 energy 0 0 0 0 0 11 Basic Principles of Color Perception Some terminology for colored light: spectral - referring to the wavelength of light the illuminant - light source power spectrum - this curve.