Color Constancy and Contextual Effects on Color Appearance
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
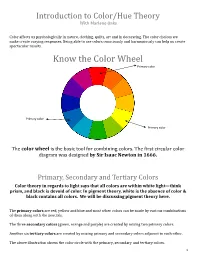
Know the Color Wheel Primary Color
Introduction to Color/Hue Theory With Marlene Oaks Color affects us psychologically in nature, clothing, quilts, art and in decorating. The color choices we make create varying responses. Being able to use colors consciously and harmoniously can help us create spectacular results. Know the Color Wheel Primary color Primary color Primary color The color wheel is the basic tool for combining colors. The first circular color diagram was designed by Sir Isaac Newton in 1666. Primary, Secondary and Tertiary Colors Color theory in regards to light says that all colors are within white light—think prism, and black is devoid of color. In pigment theory, white is the absence of color & black contains all colors. We will be discussing pigment theory here. The primary colors are red, yellow and blue and most other colors can be made by various combinations of them along with the neutrals. The three secondary colors (green, orange and purple) are created by mixing two primary colors. Another six tertiary colors are created by mixing primary and secondary colors adjacent to each other. The above illustration shows the color circle with the primary, secondary and tertiary colors. 1 Warm and cool colors The color circle can be divided into warm and cool colors. Warm colors are energizing and appear to come forward. Cool colors give an impression of calm, and appear to recede. White, black and gray are considered to be neutral. Tints - adding white to a pure hue: Terms about Shades - adding black to a pure hue: hue also known as color Tones - adding gray to a pure hue: Test for color blindness NOTE: Color theory is vast. -

Several Color Appearance Phenomena in Color Reproduction
2nd International Conference on Electronic & Mechanical Engineering and Information Technology (EMEIT-2012) Several Color Appearance Phenomena in Color Reproduction Qin-ling Dai1,a, Xiao-zhou Li2*,b, Ai Xu2 1 School of Materials Engineering (Southwest Forestry University), Kunming, China 650224 2 Key Laboratory of Pulp & Paper Science and Technology (Shandong Polytechnic University), Ministry of Education, Ji’nan, China, 250353 [email protected], bcorresponding author: [email protected] Keywords: color reproduction, color appearance, color appearance phenomena Abstract. Color perceived performance was influenced by various color appearance phenomena caused by varying viewing conditions in color reproduction process. It is necessary to do some research on the color appearance phenomena to represent the color appearance models qualitatively and quantitatively and accurate color reproduction easily. Only the phenomena were studied thoroughly, could the color transmission and reproduction be well performed. The color appearance and common color appearance phenomena of color reproduction were analyzed in this paper. And the basic theory of color appearance in color reproduction was also studied. Introduction High fidelity digital printing plays an important role in high fidelity color transmission and reproduction and it is one of the most important techniques to perform high fidelity color reproduction. High fidelity digital printing helps to perform accurate color reproduction of the original which can’t be performed because of paper and ink in traditional printing process [1]. In color printing, the color difference caused by paper, ink and viewing conditions is various. The difference is not only colorimetric difference but also different color appearance phenomena. While the different color appearance phenomenon is the leading factor to influence the color vision perceived. -

Rethinking Color Cameras
Rethinking Color Cameras Ayan Chakrabarti William T. Freeman Todd Zickler Harvard University Massachusetts Institute of Technology Harvard University Cambridge, MA Cambridge, MA Cambridge, MA [email protected] [email protected] [email protected] Abstract Digital color cameras make sub-sampled measurements of color at alternating pixel locations, and then “demo- saick” these measurements to create full color images by up-sampling. This allows traditional cameras with re- stricted processing hardware to produce color images from a single shot, but it requires blocking a majority of the in- cident light and is prone to aliasing artifacts. In this paper, we introduce a computational approach to color photogra- phy, where the sampling pattern and reconstruction process are co-designed to enhance sharpness and photographic speed. The pattern is made predominantly panchromatic, thus avoiding excessive loss of light and aliasing of high spatial-frequency intensity variations. Color is sampled at a very sparse set of locations and then propagated through- out the image with guidance from the un-aliased luminance channel. Experimental results show that this approach often leads to significant reductions in noise and aliasing arti- facts, especially in low-light conditions. Figure 1. A computational color camera. Top: Most digital color cameras use the Bayer pattern (or something like it) to sub-sample 1. Introduction color alternatingly; and then they demosaick these samples to create full-color images by up-sampling. Bottom: We propose The standard practice for one-shot digital color photog- an alternative that samples color very sparsely and is otherwise raphy is to include a color filter array in front of the sensor panchromatic. -

"He" Had Me at Blue: Color Theory and Visual
Downloaded from http://www.mitpressjournals.org/doi/pdf/10.1162/LEON_a_00677 by guest on 30 September 2021 general article “He” Had Me at Blue: Color Theory and Visual Art Barbara L. Miller a b s t r a c t Schopenhauer and Goethe argued that colors are danger- ous: When philosophers speak Blue is the colour of your yellow hair of colors, they often begin Red is the whirl of your green wheels to rant and rave. This essay addresses the confusing and ing effects. It can leave an intolera- —Kurt Schwitters treacherous history of color the- ble and “powerful impression” and ory and perception. An overview result in a type of visual incapaci- of philosophers and scientists Color Mad tation that, he suggests, “may last associated with developing for hours” [3]. Exposure to blazing theories leads into a discussion of contemporary perspectives: A friend and colleague once confided that she hated yellow light—“red” or “white” light, as the flowers: “I can’t,” she blustered, “have them in my garden.” Taussig’s notion of a “combus- fictional character cries—in real tible mixture” and “total bodily “You sound like a scene from a Hitchcock movie!” I teased, life can result in blinding after- activity” and Massumi’s idea of and Tippi Hedren as Marnie flashed before my eyes. effects; for example, walking out an “ingressive activity” are used of a dark corridor into a bright, sun- as turning points in a discussion Marnie: “First there are three taps.” of Roger Hiorns’s Seizure—an Thunder claps. Marnie swoons, wailing: “Needles . -

Spatial Filtering, Color Constancy, and the Color-Changing Dress Department of Psychology, American University, Erica L
Journal of Vision (2017) 17(3):7, 1–20 1 Spatial filtering, color constancy, and the color-changing dress Department of Psychology, American University, Erica L. Dixon Washington, DC, USA Department of Psychology and Department of Computer Arthur G. Shapiro Science, American University, Washington, DC, USA The color-changing dress is a 2015 Internet phenomenon divide among responders (as well as disagreement from in which the colors in a picture of a dress are reported as widely followed celebrity commentators) fueled a rapid blue-black by some observers and white-gold by others. spread of the photo across many online news outlets, The standard explanation is that observers make and the topic trended worldwide on Twitter under the different inferences about the lighting (is the dress in hashtag #theDress. The huge debate on the Internet shadow or bright yellow light?); based on these also sparked debate in the vision science community inferences, observers make a best guess about the about the implications of the stimulus with regard to reflectance of the dress. The assumption underlying this individual differences in color perception, which in turn explanation is that reflectance is the key to color led to a special issue of the Journal of Vision, for which constancy because reflectance alone remains invariant this article is written. under changes in lighting conditions. Here, we The predominant explanations in both scientific demonstrate an alternative type of invariance across journals (Gegenfurtner, Bloj, & Toscani, 2015; Lafer- illumination conditions: An object that appears to vary in Sousa, Hermann, & Conway, 2015; Winkler, Spill- color under blue, white, or yellow illumination does not change color in the high spatial frequency region. -

Fast Fourier Color Constancy
Fast Fourier Color Constancy Jonathan T. Barron Yun-Ta Tsai [email protected] [email protected] Abstract based techniques in computer vision, both problems reduce to just estimating the “best” illuminant from an image, and We present Fast Fourier Color Constancy (FFCC), a the question of whether that illuminant is objectively true color constancy algorithm which solves illuminant esti- or subjectively attractive is just a matter of the data used mation by reducing it to a spatial localization task on a during training. torus. By operating in the frequency domain, FFCC pro- Despite their accuracy, modern learning-based color duces lower error rates than the previous state-of-the-art by constancy algorithms are not immediately suitable as prac- 13 − 20% while being 250 − 3000× faster. This unconven- tical white balance algorithms, as practical white balance tional approach introduces challenges regarding aliasing, has several requirements besides accuracy: directional statistics, and preconditioning, which we ad- Speed - An algorithm running in a camera’s viewfinder dress. By producing a complete posterior distribution over must run at 30 FPS on mobile hardware. But a camera’s illuminants instead of a single illuminant estimate, FFCC compute budget is precious: demosaicing, face detection, enables better training techniques, an effective temporal auto exposure, etc, must also run simultaneously and in real smoothing technique, and richer methods for error analy- time. Spending more than a small fraction (say, 5 − 10%) sis. Our implementation of FFCC runs at ∼ 700 frames per of a camera’s compute budget on white balance is impracti- second on a mobile device, allowing it to be used as an ac- cal, suggesting that our speed requirement is closer to 1 − 5 curate, real-time, temporally-coherent automatic white bal- milliseconds per frame. -

Geography 222 – Color Theory in GIS Mike Pesses, Antelope Valley College
Geography 222 – Color Theory in GIS Mike Pesses, Antelope Valley College Introduction Color is a fundamental part of cartography. We have conventions that we learn early on in school; water should be blue, vegetation should be green. At the same time, we do not want to limit ourselves. While a magenta ocean may be a bit much, we can experiment with alternatives to convey a certain feeling for the map. Conventional light blue and tan world maps can feel a bit dull, whereas an “earthier” color scheme can get us thinking about exploration and piracy. A slight change in color can have major results. Color may seem like a simple enough concept, but its reproduction on paper, a television, or on a computer screen is an incredible science. To properly use color from a design standpoint, we must have at least a basic understanding of how it is produced. Color Systems Red, Green, Blue or RGB is the color system of televisions and computer screens. By simply mixing the proportions of red, green, and blue lights in screens, we can display a wealth of colors. We call this an additive system in that we add colors to make new ones. For example, if we mix red and green light, we get yellow. Mixing green and blue will produce cyan. Red and blue will make magenta. Red, green, and blue mixed together will produce white. Keep in mind that this is different from when you mixed paints in kindergarten. Mixing red, green, and blue paint will get you ‐1- Geog 222 – Color Theory in GIS, pg. -

Color Theory
color theory What is color theory? Color Theory is a set of principles used to create harmonious color combinations. Color relationships can be visually represented with a color wheel — the color spectrum wrapped onto a circle. The color wheel is a visual representation of color theory: According to color theory, harmonious color combinations use any two colors opposite each other on the color wheel, any three colors equally spaced around the color wheel forming a triangle, or any four colors forming a rectangle (actually, two pairs of colors opposite each other). The harmonious color combinations are called color schemes – sometimes the term 'color harmonies' is also used. Color schemes remain harmonious regardless of the rotation angle. Monochromatic Color Scheme The monochromatic color scheme uses variations in lightness and saturation of a single color. This scheme looks clean and elegant. Monochromatic colors go well together, producing a soothing effect. The monochromatic scheme is very easy on the eyes, especially with blue or green hues. Analogous Color Scheme The analogous color scheme uses colors that are adjacent to each other on the color wheel. One color is used as a dominant color while others are used to enrich the scheme. The analogous scheme is similar to the monochromatic, but offers more nuances. Complementary Color Scheme The complementary color scheme consists of two colors that are opposite each other on the color wheel. This scheme looks best when you place a warm color against a cool color, for example, red versus green-blue. This scheme is intrinsically high-contrast. Split Complementary Color Scheme The split complementary scheme is a variation of the standard complementary scheme. -

38 Color Constancy
38 Color Constancy DAVID H. BRAINARD AND ANA RADONJIC Vision is useful because it informs us about the physical provide a self-contained introduction, to highlight environment. In the case of colm~ two distinct functions some more recent results, and to outline what we see are generally emphasized (e.g., Jacobs, 1981; Mollon, as important challenges for the field. We focus on key 1989). First, color helps to segment objects from each concepts and avoid much in the way of technical devel other and the background. A canonical task for this opment. Other recent treatments complement the one function is locating fruit in foliage (Regan et al., 2001; provided here and provide a level of technical detail Sumner & Mollon, 2000). Second, color provides infm' beyond that introduced in this chapter (Brainard, 2009; mation about object properties (e.g., fresh fish versus Brainard & Maloney, 2011; Foster, 2011; Gilchrist, 2006; old fish; figure 38.1). Using color for this second func Kingdom, 2008; Shevell & Kingdom, 2008; Smithson, tion is enabled to the extent that perceived object color 2005; Stockman & Brainard, 2010). correlates well with object reflectance properties. Achieving such correlation is a nontrivial requirement, MEASURING CONSTANCY however, because the spectrum of the light reflected from an object confounds variation in object surface The study of constancy requires methods for measuring reflectance with variation in the illumination (figure it. The classic approach to such measurement is to 38.2; Brainard, Wandell, & Chichilnisky, 1993; Hurl assess the extent to which the color appearance of bert, 1998; Maloney, 1999). In particular, the spectrum objects varies as they are viewed under different illumi of the reflected light is the wavelength-by-wavelength nations. -

Color Blindness Fovea
Color Theory • Chapter 3b Perceiving Color • Color Constancy • Chapter 3 Perceiving Color Theory Color (so far) • Lens • Iris • Rods • Cones x3 • Fovea • Optic Nerve • Iodopsin & Rhodopsin • Chapter 3 Color Theory Perceiving Color (today) • Tri-Chromatic theory of color vision • Color Afterimage • Opponent Theory of color vision • Color Constancy • Monet’s series • Color Blindness Fovea • A tiny (1mm) area on the retina is covered with a dense collection of cones. • This region is where we see color distinctions best. Seeing Color • Still no proven model of how color perception works. • Cones contain light-sensitive pigments called Iodopsin. • Iodopsin changes when light hits it -- it is photo- sensitive. Light is effectively converted/translated into a nerve impulse…a signal to the brain. Trichromatic Theory & Opponent Theory • 19th c. Trichromatic Theory first proposed the idea of three types of cones. • Current theory -- Opponent Theory -- is that there are three types of iodopsin -- one that senses (or “sees”) red light, one that sees green, and one blue-violet. Each is “blind” to its complement. • We then combine the information from all three to perceive color. • We have mostly red- and green- sensitive cones – few blue-sensitive ones. • Diagram of suspected “wiring” of cones to ganglion Cones to Color (nerve) cells. • Light primaries are “read” individually, then results are combined. Cones to Color • Diagram of suspected trichromatic “wiring” of cones to ganglion (nerve) cells. • Light primaries are “read” individually, then results are combined. • http://www.handprint.com/HP /WCL/color1.html Opponent Processing • “As the diagram shows, the opponent processing pits the responses from one type of cone against the others. -

Middle School Science Experiment Color Theory
Middle School Science Experiment Color Theory The human eye distinguishes colors using light sensitive cells in the retina. These sensors are rods and cones. The rods give us our night vision and can function in low intensities of light, but cannot distinguish color. The cones let us see color and can resolve sharp images. The light we see, such as the light from the sun, is made up of a mixture of several colors. You will learn more about light as well as about primary and secondary colors in this experiment. Objectives In this experiment, you will: m Gain an understanding of primary and secondary colors m Learn about how a mixture of colors makes up white light m Experiment with the mixing of paint that uses pigments, not light m Take pictures of various colors and compare them when they are mixed and separated Materials m Power Macintosh G3 or better m ProScope Digital USB Microscope and software m Red, blue, and green cellophane or plastic filters m Three flashlights m Red, yellow, and blue watercolor paint m Paintbrush m Water Procedure The first activities involve light and primary colors: 1 Cover one flashlight with red cellophane, one with blue cellophane, and one with green cellophane. (You can use red, blue, and green plastic filters instead of the cellophane.) Darken the room and set up the ProScope USB microscope on the tripod pointing at a piece of white unlined paper. 2 Shine the green flashlight at the white paper. Take a picture of this image using the m0W lens. -

Chapter 7: Perceiving Color
Chapter 7: Perceiving Color -The physical dimensions of color -The psychological dimensions of color appearance (hue, saturation, brightness) -The relationship between the psychological and physical dimensions of color (Trichromacy Color opponency) - Other influences on color perception (color constancy, top-down effects) Sir Isaac Newton Newton’s Prism Experiment (1704) White light is composed of multiple colors Light Monochromatic light : one wavelength (like a laser) Physical parameters for monochromatic light 1. Wavelength 2. Intensity Heterochromatic light : many wavelengths (normal light sources) For heterochromatic light The spectral composition gives the intensity at each wavelength Monochromatic light Heterochromatic light Spectral composition of two common (heterochromatic) illuminants The spectral components of light entering the eye is the product of the illuminant and the surface reflectance of objects. Reflectance of some common surfaces and pigments 350 400 450 500 550 600 650 700 750 800 Surface reflectance of some common objects 350 400 450 500 550 600 650 700 750 800 Spectral composition of light entering the eye after being reflected from a surface = Spectral composition of the Reflectance of the surface illuminant X Consider a ripe orange illuminated with a bright monochromatic blue (420 nm) light. What color will the banana appear to be? a. bright orange b. dark/dim orange Blue (monochromatic) c. black d. dark/dim blue Relative amount of light of amount Relative Wavelength Spectral composition of light entering the