United States National Museum Bulletin 273
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Selecting Different Approaches for Palate and Pharynx Surgery
SPECIAL ISSUE 4: INVITED ARTICLE Selecting Different Approaches for Palate and Pharynx Surgery: Palatopharyngeal Arch Staging System Rodolfo Lugo-Saldaña1 , Karina Saldívar-Ponce2 , Irina González-Sáez3 , Daniela Hernández-Sirit4 , Patricia Mireles-García5 ABSTRACT The examination of the anatomical structures involved in the upper airway collapse in patients with the obstructive sleep apnea-hypopnea syndrome (OSAHS) is a key for integrated evaluation of patients. Our proposal is for a noninvasive classification system that guides us about the presence of anatomical differences between the palatopharyngeal muscle (PFM). The functions of the PFM are narrowing the isthmus, descending the palate, and raising the larynx during swallowing; these characteristics give the PFM a special role in the collapse of the lateral pharyngeal wall. Complete knowledge of the anatomy and classification of different variants can guide us to choose the appropriate surgical procedures for the lateral wall collapse. Until now there is not a consensus about description of the trajectory or anatomical variants of the PFM into oropharynx, the distance between both muscles, and the muscle tone. Here we also present the relationship between the lateral wall surgeries currently available (lateral pharyngoplasty by Cahali, expansion sphincteroplasty by Pang, relocation pharyngoplasty by Li, Roman blinds pharyngoplasty by Mantovani, and barbed sutures pharyngoplasty by Vicini) with the proposed classification of the palatopharyngeal arch staging system (PASS). Keywords: -

Iliopsoas Tendonitis/Bursitis Exercises
ILIOPSOAS TENDONITIS / BURSITIS What is the Iliopsoas and Bursa? The iliopsoas is a muscle that runs from your lower back through the pelvis to attach to a small bump (the lesser trochanter) on the top portion of the thighbone near your groin. This muscle has the important job of helping to bend the hip—it helps you to lift your leg when going up and down stairs or to start getting out of a car. A fluid-filled sac (bursa) helps to protect and allow the tendon to glide during these movements. The iliopsoas tendon can become inflamed or overworked during repetitive activities. The tendon can also become irritated after hip replacement surgery. Signs and Symptoms Iliopsoas issues may feel like “a pulled groin muscle”. The main symptom is usually a catch during certain movements such as when trying to put on socks or rising from a seated position. You may find yourself leading with your other leg when going up the stairs to avoid lifting the painful leg. The pain may extend from the groin to the inside of the thigh area. Snapping or clicking within the front of the hip can also be experienced. Do not worry this is not your hip trying to pop out of socket but it is usually the iliopsoas tendon rubbing over the hip joint or pelvis. Treatment Conservative treatment in the form of stretching and strengthening usually helps with the majority of patients with iliopsoas bursitis. This issue is the result of soft tissue inflammation, therefore rest, ice, anti- inflammatory medications, physical therapy exercises, and/or injections are effective treatment options. -

14-Anatomy of Forearm
FOREARM By : Prof.Saeed Abulmakarem. Dr. Sanaa Al-Sharawy OBJECTIVES §At the end of this lecture, the student should able to : §List the names of the Flexors Group of Forearm (superficial & deep muscles). §Identify the common flexor origin of flexor muscles and their innervation & movements. §Identify supination & poronation and list the muscles produced these 2 movements. §List the names of the Extensor Group of Forearm (superficial & deep muscles). §Identify the common extensor origin of extensor musles and their innervation & movements. n The forearm extends from elbow to wrist. n It posses two bones radius laterally & Ulna medially. n The two bones are connected together by the interosseous membrane. n This membrane allows movement of Pronation and Supination while the two bones are connected together. n Also it gives origin for the deep muscles. § The forearm is Fascial Compartments of the Forearm enclosed in a sheath of deep fascia, which is attached to the posterior border of the ulna . §This fascial sheath, together with the interosseous membrane & fibrous intermuscular septa, divides the forearm into compartments, each having its own muscles, nerves, and blood supply. These muscles: 8 FLEXOR GROUP § Act on the elbow & wrist joints and those of the fingers. § Form fleshy masses in the proximal part and become tendinous in the distal part of the forearm. •Arranged in three groups: I-Superficial: 4 Ø Pronator teres Ø Flexor carpi radialis Ø Palmaris longus III- Deep: 3 Ø Flexor carpi ulnaris Ø Flexor digitorum profundus II-Intermediate: 1 Ø Flexor pollicis longus Ø Ø Flexor digitorum superficialis Pronator quadratus n Superficial Flexors: n They arise - more or less- from the common flexor origin (front of medial epicondyle). -

The Effect of Stretching Hamstring, Gastrocnemius, Iliopsoas
Open Journal of Therapy and Rehabilitation, 2015, 3, 139-145 Published Online November 2015 in SciRes. http://www.scirp.org/journal/ojtr http://dx.doi.org/10.4236/ojtr.2015.34019 The Effect of Stretching Hamstring, Gastrocnemius, Iliopsoas and Back Muscles on Pain and Functional Activities in Patients with Chronic Low Back Pain: A Randomized Clinical Trial Hamada E. Seif1, Aqeel Alenazi2, Sahar Mahmoud Hassan1, Shaji John Kachanathu3, Ashraf R. Hafez1* 1Cairo UniversityHospital, Cairo University, Cairo, Egypt 2Physical Therapy and Rehabilitation Department, College of Applied Medical Sciences, Salman Bin Abdulaziz University, Alkharj, Saudi Arabia 3Collage of Applied Medical Sciences, King Saud University, Riyadh, Saudi Arabia Received 15 September 2015; accepted 6 November 2015; published 9 November 2015 Copyright © 2015 by authors and Scientific Research Publishing Inc. This work is licensed under the Creative Commons Attribution International License (CC BY). http://creativecommons.org/licenses/by/4.0/ Abstract A back pain lasting more than 12 weeks has been defined as a chronic low back pain (LBP) [1]. More than half of people suffer from LBP [1]. The purpose of this study was to examine the effect of gastrocnemius muscle stretching in the treatment of chronic low back pain. Methods: Forty pa- tients with chronic low back pain, ages ranging from 25 to 40 years, were recruited and divided randomly into two groups. The control group followed a physical therapy program that included stretching exercises for back, hamstring and iliopsoas muscles. Strengthening exercises for abdo- minal muscle and postural instructions for activities of daily living were also performed. The ex- perimental group followed the same control-group exercises with the addition of stretching exer- cises for gastrocnemius muscles. -

Gross Anatomy
www.BookOfLinks.com THE BIG PICTURE GROSS ANATOMY www.BookOfLinks.com Notice Medicine is an ever-changing science. As new research and clinical experience broaden our knowledge, changes in treatment and drug therapy are required. The authors and the publisher of this work have checked with sources believed to be reliable in their efforts to provide information that is complete and generally in accord with the standards accepted at the time of publication. However, in view of the possibility of human error or changes in medical sciences, neither the authors nor the publisher nor any other party who has been involved in the preparation or publication of this work warrants that the information contained herein is in every respect accurate or complete, and they disclaim all responsibility for any errors or omissions or for the results obtained from use of the information contained in this work. Readers are encouraged to confirm the infor- mation contained herein with other sources. For example and in particular, readers are advised to check the product information sheet included in the package of each drug they plan to administer to be certain that the information contained in this work is accurate and that changes have not been made in the recommended dose or in the contraindications for administration. This recommendation is of particular importance in connection with new or infrequently used drugs. www.BookOfLinks.com THE BIG PICTURE GROSS ANATOMY David A. Morton, PhD Associate Professor Anatomy Director Department of Neurobiology and Anatomy University of Utah School of Medicine Salt Lake City, Utah K. Bo Foreman, PhD, PT Assistant Professor Anatomy Director University of Utah College of Health Salt Lake City, Utah Kurt H. -

Take Care of Your Feet .Pub
Newsletter Feb / Mar 2018 www.chiropractorcapalaba.websyte.com.au Publisher Dr. Patrick J. Shwaluk Educated - Safe - Effective Doctor of Chiropractic Spine Care Certified Chiropractic Sports Physician Many patients consider this Take care of your feet Feet are complicated. Immobilizing feet by newsletter as a squeezing them into tight fitting reminder to come in shoes and standing on them all for their monthly day can causes the muscles to good spinal health shorten and cramp. When the check up. Now is a arches drop the joints, fascia, tendons and ligaments to get over good time to book stretched, thicken with scar tissue, your “tune up” stiffen, become brittle and painful. appointment . By the time someone experiences plantar fasciitis they have a drop in one or more of the arches, stiffening Figure 2: Plantar Facia of some of the joints of the feet Clinic Hours and a 50% thickening Figure 1: 3 arches The stretches in figure 3 should be held for of the plantar fascia three minutes each. Long duration stretches, - Mon 10am 7pm ligament, figure 2. referred to as “melting stretches”, are required - Tues 9 am 12pm to cause plastic deformation of the fascia and - Wed 10am 6pm Each foot has 26 bones and 3 arches, an increase in its length. Short duration - Thurs 3pm 7pm figure 1 . The transverse arch, A-B, is stretches will feel good and mobilize the feet - Fri 9am 4pm associated with metatarsalgia and Morton’s but not change the overall length of the fascia. - Sat 9:30 am 12:30pm neuromas. The lateral longitudinal arch, B-C, is associated with lateral foot pain. -

Lower Limb Osteofascial Compartments
Lower limb Osteofascial compartments Zdenek Halata, Department of Anatomy, First Faculty of Medicine, Charles University, Prague Pelvis – osteofascial compartments: 1. Lacuna vasorum et Lacuna musculorum: between lig. inguinale (spina iliaca ant. sup. and tuberculum pubicum) and os ilium and ramus superior ossis pubis a) Lacuna vasorum – A.iliaca externa – A. femoralis V.iliaca externa – V. femoralis b) Lacuna musculorum M. iliopsoas N. femoralis N. cutaneus femoris lateralis 2. Canalis obturatorius Vasa obturatoria et N. obturatorius 3. Foramen suprapiriforme Vasa glutea sup., N. gluteus superior 4. Foramen infrapiriforme Vasa glutea inf., N.gluteus inf., N.cutaneus femoris post., N. ischiadicus Thigh – osteofascial compartments: 1. compartment of adductor muscles: m. adductor magnus, longus and brevis m. gracilis with separate compartment 1. compartment of extensor muscles: m.quadriceps femoris, m.sartorius with separate compartment 3. compartment of flexor muscles: m.semimebranosus, m.semitendinosus m.biceps femoris Septum intermusculare femoris laterale Septum intermusculare femoris mediale: with a. and v. femoralis and N. saphenus Septum intermusculare femoris posterius: with n.ischiadicus, aa.perforantes and vv.perforantes 4. Canalis vastoadductorius: between fossa iliopectinea and fossa poplitea Calf - osteofascial compartments Compartment of extensor muscles: m.tibialis anterior, m.extensor hallucis longus, m.extensor digitorum longus Compartment of superficial flexor muscles: m.soleus, m.gastrocnemius Compartment of deep flexor muscles: m.tibialis posterior, m.flexor hallucis longus, m.flexor digitorum longus Compartment of peroneus muscles: m.peroneus longus, m.peroneus brevis Fascia cruris with superficial and deep layer (lamina superficialis and profunda) Membrana interossea cruris Septum intermusculare cruris anterius Septum intermusculare cruris posterius Foot - osteofascial compartments Compartment with extensor tendons in synovial sheaths Compartment with flexors for the 5. -

Head & Neck Surgery Course
Head & Neck Surgery Course Parapharyngeal space: surgical anatomy Dr Pierfrancesco PELLICCIA Pr Benjamin LALLEMANT Service ORL et CMF CHU de Nîmes CH de Arles Introduction • Potential deep neck space • Shaped as an inverted pyramid • Base of the pyramid: skull base • Apex of the pyramid: greater cornu of the hyoid bone Introduction • 2 compartments – Prestyloid – Poststyloid Anatomy: boundaries • Superior: small portion of temporal bone • Inferior: junction of the posterior belly of the digastric and the hyoid bone Anatomy: boundaries Anatomy: boundaries • Posterior: deep fascia and paravertebral muscle • Anterior: pterygomandibular raphe and medial pterygoid muscle fascia Anatomy: boundaries • Medial: pharynx (pharyngobasilar fascia, pharyngeal wall, buccopharyngeal fascia) • Lateral: superficial layer of deep fascia • Medial pterygoid muscle fascia • Mandibular ramus • Retromandibular portion of the deep lobe of the parotid gland • Posterior belly of digastric muscle • 2 ligaments – Sphenomandibular ligament – Stylomandibular ligament Aponeurosis and ligaments Aponeurosis and ligaments • Stylopharyngeal aponeurosis: separates parapharyngeal spaces to two compartments: – Prestyloid – Poststyloid • Cloison sagittale: separates parapharyngeal and retropharyngeal space Aponeurosis and ligaments Stylopharyngeal aponeurosis Muscles stylohyoidien Stylopharyngeal , And styloglossus muscles Prestyloid compartment Contents: – Retromandibular portion of the deep lobe of the parotid gland – Minor or ectopic salivary gland – CN V branch to tensor -

Compiled for Lower Limb
Updated: December, 9th, 2020 MSI ANATOMY LAB: STRUCTURE LIST Lower Extremity Lower Extremity Osteology Hip bone Tibia • Greater sciatic notch • Medial condyle • Lesser sciatic notch • Lateral condyle • Obturator foramen • Tibial plateau • Acetabulum o Medial tibial plateau o Lunate surface o Lateral tibial plateau o Acetabular notch o Intercondylar eminence • Ischiopubic ramus o Anterior intercondylar area o Posterior intercondylar area Pubic bone (pubis) • Pectineal line • Tibial tuberosity • Pubic tubercle • Medial malleolus • Body • Superior pubic ramus Patella • Inferior pubic ramus Fibula Ischium • Head • Body • Neck • Ramus • Lateral malleolus • Ischial tuberosity • Ischial spine Foot • Calcaneus Ilium o Calcaneal tuberosity • Iliac fossa o Sustentaculum tali (talar shelf) • Anterior superior iliac spine • Anterior inferior iliac spine • Talus o Head • Posterior superior iliac spine o Neck • Posterior inferior iliac spine • Arcuate line • Navicular • Iliac crest • Cuboid • Body • Cuneiforms: medial, intermediate, and lateral Femur • Metatarsals 1-5 • Greater trochanter • Phalanges 1-5 • Lesser trochanter o Proximal • Head o Middle • Neck o Distal • Linea aspera • L • Lateral condyle • L • Intercondylar fossa (notch) • L • Medial condyle • L • Lateral epicondyle • L • Medial epicondyle • L • Adductor tubercle • L • L • L • L • 1 Updated: December, 9th, 2020 Lab 3: Anterior and Medial Thigh Anterior Thigh Medial thigh General Structures Muscles • Fascia lata • Adductor longus m. • Anterior compartment • Adductor brevis m. • Medial compartment • Adductor magnus m. • Great saphenous vein o Adductor hiatus • Femoral sheath o Compartments and contents • Pectineus m. o Femoral canal and ring • Gracilis m. Muscles & Associated Tendons Nerves • Tensor fasciae lata • Obturator nerve • Iliotibial tract (band) • Femoral triangle: Boundaries Vessels o Inguinal ligament • Obturator artery o Sartorius m. • Femoral artery o Adductor longus m. -
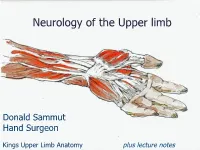
Neurology of the Upper Limb
Neurology of the Upper limb Donald Sammut Hand Surgeon Kings Upper Limb Anatomy plus lecture notes The$Neck$ The$Nerve$roots$which$supply$the$Upper$Limb$are$C5$to$T1$ Pre<fixed$(C4$to$C8)$and$Post<fixed$(C6$to$T2)$plexus$not$uncommon.$ Also$common$contributions$from$C4$and$from$T2$in$a$normally$rooted$plexus.$ $ The$anterior$nerve$roots$emerge$between$the$vertebrae$and$immediately$pass$ $through$the$first$area$of$possible$compression:$ The$root$nerve$canal$is$bounded$$ Anteriorly$by$the$posterior$margin$of$the$intervertebral$disc$and$$ Posteriorly,$by$the$facet$joint$between$vertebrae.$ $ Pathology$of$the$disc,$or$joint,$or$both,$can$narrow$this$channel$and$compress$ $the$nerve$root$ The$roots$emerge$from$the$cervical$spine$into$the$plane$between$$ Scalenius$Anterior$and$Scalenius$Medius.$$ $ Scalenius*Anterior:** Origin:$Anterior$tubercles$of$Cervical$vertebae$C3$to$6$(C6$tubercle$is$the$Carotid$tubercle)$ Insertion:$The$scalene$tubercle$on$inner$border/upper$surface$1st$rib$ $ Scalenius*Medius:* Origin:$Posterior$tubercles$of$all$cervical$vertebrae$ Insertion:$Quadrangular$area$between$the$neck$and$subclavian$groove$1st$rib$ $ Exiting$from$the$Scalenes,$the$trunks$lie$in$the$posterior$triangle$of$the$neck.$ The$posterior$triangle$is$bounded$anteriorly$by$SternoCleidoMastoid$and$$ posteriorly$by$the$Trapezius.$ The$inferior$border$is$the$clavicle$.$ The$apex$of$the$triangle$superiorly$is$at$the$back$of$the$skull$on$the$superior$nuchal$line$ $ $ The$Posterior$Triangle$ SternoCleidoMastoid$ Trapezius$ Scalenius$Medius$ Scalenius$Anterior$ -

Deep Neck Infections 55
Deep Neck Infections 55 Behrad B. Aynehchi Gady Har-El Deep neck space infections (DNSIs) are a relatively penetrating trauma, surgical instrument trauma, spread infrequent entity in the postpenicillin era. Their occur- from superfi cial infections, necrotic malignant nodes, rence, however, poses considerable challenges in diagnosis mastoiditis with resultant Bezold abscess, and unknown and treatment and they may result in potentially serious causes (3–5). In inner cities, where intravenous drug or even fatal complications in the absence of timely rec- abuse (IVDA) is more common, there is a higher preva- ognition. The advent of antibiotics has led to a continu- lence of infections of the jugular vein and carotid sheath ing evolution in etiology, presentation, clinical course, and from contaminated needles (6–8). The emerging practice antimicrobial resistance patterns. These trends combined of “shotgunning” crack cocaine has been associated with with the complex anatomy of the head and neck under- retropharyngeal abscesses as well (9). These purulent col- score the importance of clinical suspicion and thorough lections from direct inoculation, however, seem to have a diagnostic evaluation. Proper management of a recog- more benign clinical course compared to those spreading nized DNSI begins with securing the airway. Despite recent from infl amed tissue (10). Congenital anomalies includ- advances in imaging and conservative medical manage- ing thyroglossal duct cysts and branchial cleft anomalies ment, surgical drainage remains a mainstay in the treat- must also be considered, particularly in cases where no ment in many cases. apparent source can be readily identifi ed. Regardless of the etiology, infection and infl ammation can spread through- Q1 ETIOLOGY out the various regions via arteries, veins, lymphatics, or direct extension along fascial planes. -

Brachial Plexus and Branches Axillary A
Regional Anatomy of the Upper Limb 1 Wu Genghua Medical College Yangzhou University Anterior surface of the •Incisions: Upper Limb Make the skin incisions indicated in figure and reflect the skin flaps. Be careful not to cut too deep. As you remove the skin, look for the cephalic v., basilic v. and median cubital v. Superficial veins and nerves Mamma Structures Contains skin, mammary glands and adipose tissue Consists of 15 to 20 Lobes of mammary gland that radiate outward from the nipple lactiferous duct lactiferous sinuse Suspensory ligaments of breast (cooper’s ligaments): connective tissue septa that extend from the skin to the deep fascia Pectoral Region Superficial structures Superficial n. Supraclavicular n. Anterior cutaneous branches of intercostal n. lateral cutaneous branches of intercostal n. Muscles connecting the upper limb to the thoracic wall Pectoralis major 胸大肌 Subclevius 锁骨下肌 Pectoralis minor 胸小肌 Serratus anterior 前锯肌 Pectoral Region Deep structures clavipectoral fascia The deep fascia which extends between subclavius, coracoid process and pectoralis minor muscles The structures pass through the clavipectoral fascia . Cephalic v. Thoracoacromial a. Lateral pectoral n. Axillary region 腋区 Boundaries of the axillary fossa Apex is bounded by Middle 1/3 of clavicle Lateral border of first rib Upper border of the scapula Base is formed by the skin stretching between the anterior and posterior walls Anterior wall Formed by pectoralis major, pectoralis minor and subclavius muscles Boundaries of the axillary fossa The posterior wall latissimus dorsi teres major subscapularis and scapula Trilateral and quadrilateral space The posterior humeral circumflex a. and axillary n. pass through the quadrilateral foramen.