Leaf Herbivory by Insects During Summer Reduces Overwinter Browsing by Moose Brian P
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Willows of Interior Alaska
1 Willows of Interior Alaska Dominique M. Collet US Fish and Wildlife Service 2004 2 Willows of Interior Alaska Acknowledgements The development of this willow guide has been made possible thanks to funding from the U.S. Fish and Wildlife Service- Yukon Flats National Wildlife Refuge - order 70181-12-M692. Funding for printing was made available through a collaborative partnership of Natural Resources, U.S. Army Alaska, Department of Defense; Pacific North- west Research Station, U.S. Forest Service, Department of Agriculture; National Park Service, and Fairbanks Fish and Wildlife Field Office, U.S. Fish and Wildlife Service, Department of the Interior; and Bonanza Creek Long Term Ecological Research Program, University of Alaska Fairbanks. The data for the distribution maps were provided by George Argus, Al Batten, Garry Davies, Rob deVelice, and Carolyn Parker. Carol Griswold, George Argus, Les Viereck and Delia Person provided much improvement to the manuscript by their careful editing and suggestions. I want to thank Delia Person, of the Yukon Flats National Wildlife Refuge, for initiating and following through with the development and printing of this guide. Most of all, I am especially grateful to Pamela Houston whose support made the writing of this guide possible. Any errors or omissions are solely the responsibility of the author. Disclaimer This publication is designed to provide accurate information on willows from interior Alaska. If expert knowledge is required, services of an experienced botanist should be sought. Contents -

Poplars and Willows: Trees for Society and the Environment / Edited by J.G
Poplars and Willows Trees for Society and the Environment This volume is respectfully dedicated to the memory of Victor Steenackers. Vic, as he was known to his friends, was born in Weelde, Belgium, in 1928. His life was devoted to his family – his wife, Joanna, his 9 children and his 23 grandchildren. His career was devoted to the study and improve- ment of poplars, particularly through poplar breeding. As Director of the Poplar Research Institute at Geraardsbergen, Belgium, he pursued a lifelong scientific interest in poplars and encouraged others to share his passion. As a member of the Executive Committee of the International Poplar Commission for many years, and as its Chair from 1988 to 2000, he was a much-loved mentor and powerful advocate, spreading scientific knowledge of poplars and willows worldwide throughout the many member countries of the IPC. This book is in many ways part of the legacy of Vic Steenackers, many of its contributing authors having learned from his guidance and dedication. Vic Steenackers passed away at Aalst, Belgium, in August 2010, but his work is carried on by others, including mem- bers of his family. Poplars and Willows Trees for Society and the Environment Edited by J.G. Isebrands Environmental Forestry Consultants LLC, New London, Wisconsin, USA and J. Richardson Poplar Council of Canada, Ottawa, Ontario, Canada Published by The Food and Agriculture Organization of the United Nations and CABI CABI is a trading name of CAB International CABI CABI Nosworthy Way 38 Chauncey Street Wallingford Suite 1002 Oxfordshire OX10 8DE Boston, MA 02111 UK USA Tel: +44 (0)1491 832111 Tel: +1 800 552 3083 (toll free) Fax: +44 (0)1491 833508 Tel: +1 (0)617 395 4051 E-mail: [email protected] E-mail: [email protected] Website: www.cabi.org © FAO, 2014 FAO encourages the use, reproduction and dissemination of material in this information product. -

Evaluating DNA Barcoding for Species Identification And
Evaluating DNA Barcoding for Species Identification and Discovery in European Gracillariid Moths Carlos Lopez-Vaamonde, Natalia Kirichenko, Alain Cama, Camiel Doorenweerd, H. Charles J. Godfray, Antoine Guiguet, Stanislav Gomboc, Peter Huemer, Jean-François Landry, Ales Laštůvka, et al. To cite this version: Carlos Lopez-Vaamonde, Natalia Kirichenko, Alain Cama, Camiel Doorenweerd, H. Charles J. Godfray, et al.. Evaluating DNA Barcoding for Species Identification and Discovery in Euro- pean Gracillariid Moths. Frontiers in Ecology and Evolution, Frontiers Media S.A, 2021, 9, 10.3389/fevo.2021.626752. hal-03149644 HAL Id: hal-03149644 https://hal.sorbonne-universite.fr/hal-03149644 Submitted on 23 Feb 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. fevo-09-626752 February 14, 2021 Time: 16:51 # 1 ORIGINAL RESEARCH published: 18 February 2021 doi: 10.3389/fevo.2021.626752 Evaluating DNA Barcoding for Species Identification and Discovery in European Gracillariid Moths Carlos Lopez-Vaamonde1,2*, Natalia Kirichenko3,4, Alain Cama5, Camiel Doorenweerd6,7, H. Charles J. Godfray8, Antoine Guiguet9, Stanislav Gomboc10, Peter Huemer11, Jean-François Landry12, Ales Lašt ˚uvka13, Zdenek Lašt ˚uvka14, Kyung Min Lee15, David C. Lees16, Marko Mutanen15, Erik J. -
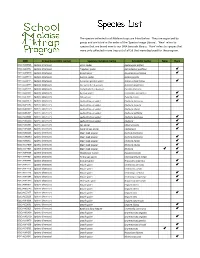
Species List
The species collected in all Malaise traps are listed below. They are organized by group and are listed in the order of the 'Species Image Library'. ‘New’ refers to species that are brand new to our DNA barcode library. 'Rare' refers to species that were only collected in one trap out of all 59 that were deployed for the program. -

Forests of Southeast and South-Central Alaska, 2004-2008
Forests of Southeast and South-Central Alaska, 2004–2008 Five-Year Forest Inventory and Analysis Report General Technical Report United States Forest Pacific Northwest PNW-GTR-835 Department of D E E P R A U Service Research Station R T LT March 2011 Agriculture MENT OF AGRICU The Forest Service of the U.S. Department of Agriculture is dedicated to the principle of multiple use management of the Nation’s forest resources for sustained yields of wood, water, forage, wildlife, and recreation. Through forestry research, cooperation with the States and private forest owners, and management of the national forests and national grasslands, it strives—as directed by Congress—to provide increasingly greater service to a growing Nation. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable, sex, marital status, familial status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, Room 1400 Independence Avenue, SW, Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer. Technical Editors Tara M. -

Redalyc.Interactions Among Host Plants, Lepidoptera Leaf Miners And
SHILAP Revista de Lepidopterología ISSN: 0300-5267 [email protected] Sociedad Hispano-Luso-Americana de Lepidopterología España Yefremova, Z. A.; Kravchenko, V. D. Interactions among host plants, Lepidoptera leaf miners and their parasitoids in the forest- steppe zone of Russia (Insecta: Lepidoptera, Hymenoptera) SHILAP Revista de Lepidopterología, vol. 43, núm. 170, junio, 2015, pp. 271-280 Sociedad Hispano-Luso-Americana de Lepidopterología Madrid, España Available in: http://www.redalyc.org/articulo.oa?id=45541421012 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative 271-280 Interactions among host 3/6/15 10:45 Página 271 SHILAP Revta. lepid., 43 (170), junio 2015: 271-280 eISSN: 2340-4078 ISSN: 0300-5267 Interactions among host plants, Lepidoptera leaf miners and their parasitoids in the forest-steppe zone of Russia (Insecta: Lepidoptera, Hymenoptera) Z. A. Yefremova & V. D. Kravchenko Abstract The article reports on the quantitative description of the food web structure of the community consisting of 65 species of Lepidoptera leaf miners reared from 34 plant species, as well as 107 species of parasitoid eulophid wasps (Hymenoptera: Eulophidae). The study was conducted in the forest-steppe zone of the Middle Volga in Russia over 13 years (2000-2012). Leaf miners have been found to be highly host plant-specific. Most of them are associated with only one or two plant species and therefore the number of links between trophic levels is 73, which is close to the total number of Lepidoptera species (linkage density is 1.12). -

Leafmining Insects Fact Sheet No
Leafmining Insects Fact Sheet No. 5.548 Insect Series|Trees and Shrubs by W.S. Cranshaw, D.A. Leatherman and J.R. Feucht* Leafminers are insects that have a habit of and/or its droppings (frass). Leaf spotting Quick Facts feeding within leaves or needles, producing fungi cause these areas to collapse, without tunneling injuries. Several kinds of insects any tunneling. • Leafminers are insects have developed this habit, including larvae of that feed within a leaf, moths (Lepidoptera), beetles (Coleoptera), producing large blotches or sawflies (Hymenoptera) and flies (Diptera). Common Leafminers meandering tunnels. Most of these insects feed for their entire of Trees and Shrubs • Although leafminer injuries larval period within the leaf. Some will also Sawfly Leafminers. Most sawflies chew are conspicuous, most pupate within the leaf mine, while others on the surface of leaves, but four species have larvae that cut their way out when full- found in Colorado develop as leafminers of leafminers produce injuries grown to pupate in the soil. woody plants. Adults are small, dark-colored, that have little, if any, effect on Leafminers are sometimes classified by non-stinging wasps that insert eggs into the plant health. the pattern of the mine which they create. newly formed leaves. The developing larvae • Most leafminers have many Serpentine leaf mines wind snake-like across produce large blotch mines in leaves during natural controls that will the leaf gradually widening as the insect late spring. The sawfly leafminers produced a normally provide good control grows. More common are various blotch single generation each year. leaf mines which are generally irregularly Elm leafminer (Kaliofenusa ulmi) is of leafminers. -

Forest Health Conditions in Alaska 2020
Forest Service U.S. DEPARTMENT OF AGRICULTURE Alaska Region | R10-PR-046 | April 2021 Forest Health Conditions in Alaska - 2020 A Forest Health Protection Report U.S. Department of Agriculture, Forest Service, State & Private Forestry, Alaska Region Karl Dalla Rosa, Acting Director for State & Private Forestry, 1220 SW Third Avenue, Portland, OR 97204, [email protected] Michael Shephard, Deputy Director State & Private Forestry, 161 East 1st Avenue, Door 8, Anchorage, AK 99501, [email protected] Jason Anderson, Acting Deputy Director State & Private Forestry, 161 East 1st Avenue, Door 8, Anchorage, AK 99501, [email protected] Alaska Forest Health Specialists Forest Service, Forest Health Protection, http://www.fs.fed.us/r10/spf/fhp/ Anchorage, Southcentral Field Office 161 East 1st Avenue, Door 8, Anchorage, AK 99501 Phone: (907) 743-9451 Fax: (907) 743-9479 Betty Charnon, Invasive Plants, FHM, Pesticides, [email protected]; Jessie Moan, Entomologist, [email protected]; Steve Swenson, Biological Science Technician, [email protected] Fairbanks, Interior Field Office 3700 Airport Way, Fairbanks, AK 99709 Phone: (907) 451-2799, Fax: (907) 451-2690 Sydney Brannoch, Entomologist, [email protected]; Garret Dubois, Biological Science Technician, [email protected]; Lori Winton, Plant Pathologist, [email protected] Juneau, Southeast Field Office 11175 Auke Lake Way, Juneau, AK 99801 Phone: (907) 586-8811; Fax: (907) 586-7848 Isaac Dell, Biological Scientist, [email protected]; Elizabeth Graham, Entomologist, [email protected]; Karen Hutten, Aerial Survey Program Manager, [email protected]; Robin Mulvey, Plant Pathologist, [email protected] State of Alaska, Department of Natural Resources Division of Forestry 550 W 7th Avenue, Suite 1450, Anchorage, AK 99501 Phone: (907) 269-8460; Fax: (907) 269-8931 Jason Moan, Forest Health Program Coordinator, [email protected]; Martin Schoofs, Forest Health Forester, [email protected] University of Alaska Fairbanks Cooperative Extension Service 219 E. -

Redalyc.New Records of Lepidoptera from the Iberian Peninsula for 2015
SHILAP Revista de Lepidopterología ISSN: 0300-5267 [email protected] Sociedad Hispano-Luso-Americana de Lepidopterología España Lastuvka, A.; Lastuvka, Z. New records of Lepidoptera from the Iberian Peninsula for 2015 (Insecta: Lepidoptera) SHILAP Revista de Lepidopterología, vol. 43, núm. 172, diciembre, 2015, pp. 633-644 Sociedad Hispano-Luso-Americana de Lepidopterología Madrid, España Available in: http://www.redalyc.org/articulo.oa?id=45543699008 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative SHILAP Revta. lepid., 43 (172), diciembre 2015: 633-644 eISSN: 2340-4078 ISSN: 0300-5267 New records of Lepidoptera from the Iberian Peninsula for 2015 (Insecta: Lepidoptera) A. Lastuvka & Z. Lastuvka Abstract New records of Nepticulidae, Heliozelidae, Adelidae, Tischeriidae, Gracillariidae, Argyresthiidae, Lyonetiidae and Sesiidae for Portugal and Spain are presented. Stigmella minusculella (Herrich-Schäffer, 1855), S. tormentillella (Herrich-Schäffer, 1860), Parafomoria helianthemella (Herrich-Schäffer, 1860), Antispila metallella ([Denis & Schiffermüller], 1775), Nematopogon metaxella (Hübner, [1813]), Tischeria dodonaea Stainton, 1858, Coptotriche gaunacella (Duponchel, 1843), Caloptilia fidella (Reutti, 1853), Phyllonorycter monspessulanella (Fuchs, 1897), P. spinicolella (Zeller, 1846), Lyonetia prunifoliella -

Mining Patterns of the Aspen Leaf Miner, Phyllocnistis Populiella, on Its Host Plant, Populus Tremuloides
Western North American Naturalist Volume 71 Number 1 Article 5 4-20-2011 Mining patterns of the aspen leaf miner, Phyllocnistis populiella, on its host plant, Populus tremuloides Martin L. Morton University of Tulsa, Tulsa, Oklahoma, [email protected] Maria E. Pereyra University of Tulsa, Tulsa, Oklahoma, [email protected] Follow this and additional works at: https://scholarsarchive.byu.edu/wnan Part of the Anatomy Commons, Botany Commons, Physiology Commons, and the Zoology Commons Recommended Citation Morton, Martin L. and Pereyra, Maria E. (2011) "Mining patterns of the aspen leaf miner, Phyllocnistis populiella, on its host plant, Populus tremuloides," Western North American Naturalist: Vol. 71 : No. 1 , Article 5. Available at: https://scholarsarchive.byu.edu/wnan/vol71/iss1/5 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Western North American Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Western North American Naturalist 71(1), © 2011, pp. 33–37 MINING PATTERNS OF THE ASPEN LEAF MINER, PHYLLOCNISTIS POPULIELLA, ON ITS HOST PLANT, POPULUS TREMULOIDES Martin L. Morton1,2 and Maria E. Pereyra1 ABSTRACT.—Feeding behavior of aspen leaf miner (Phyllocnistis populiella) larvae on quaking aspen (Populus tremu- loides) leaves was examined, by using image-analysis software, to determine the pattern and distance of the larvae’s mining trails and measure the surface area of infested leaves and consumed leaf tissue. Mining pathways between hatching and pupation sites were serpentine and generally followed predictable, sequential changes in orientation. -

Lepidoptera: Gracillariidae
Document generated on 10/01/2021 6:31 p.m. Phytoprotection --> See the erratum for this article Chalcidoid parasitoids of Micrurapteryx sophorivora [Lepidoptera: Gracillariidae] in Kuluncak, Turkey Parasitoïdes chalcidoïdes de Micrurapteryx sophorivora [Lepidoptera : Gracillariidae] de la région de Kuluncak en Turquie Lütfiye Gençer and Selma Seven Volume 86, Number 2, août 2005 Article abstract This study deals with the parasitoids of Micrurapteryx sophorivora. Parasitoids URI: https://id.erudit.org/iderudit/012513ar of M. sophorivora were investigated on Robinia pseudoacacia in 2004 in the DOI: https://doi.org/10.7202/012513ar Kuluncak district, Turkey. Seven parasitoid species, Baryscapus nigroviolaceus, Cirrospilus pictus, Necremnus croton, Neochrysocharis arvensis, See table of contents Neochrysocharis formosa, Pnigalio sp. and Pteromalus sp., were reared. Necremnus croton was found to be the most common parasitoid. All the parasitoids reared were recorded for the first time from Micrurapteryx Publisher(s) sophorivora. Société de protection des plantes du Québec (SPPQ) ISSN 0031-9511 (print) 1710-1603 (digital) Explore this journal Cite this article Gençer, L. & Seven, S. (2005). Chalcidoid parasitoids of Micrurapteryx sophorivora [Lepidoptera: Gracillariidae] in Kuluncak, Turkey / Parasitoïdes chalcidoïdes de Micrurapteryx sophorivora [Lepidoptera : Gracillariidae] de la région de Kuluncak en Turquie. Phytoprotection, 86(2), 133–134. https://doi.org/10.7202/012513ar Tous droits réservés © La société de protection des plantes du Québec, 2005 This document is protected by copyright law. Use of the services of Érudit (including reproduction) is subject to its terms and conditions, which can be viewed online. https://apropos.erudit.org/en/users/policy-on-use/ This article is disseminated and preserved by Érudit. Érudit is a non-profit inter-university consortium of the Université de Montréal, Université Laval, and the Université du Québec à Montréal. -

Issue Information
Systematic Entomology (2017), 42, 60–81 DOI: 10.1111/syen.12210 A molecular phylogeny and revised higher-level classification for the leaf-mining moth family Gracillariidae and its implications for larval host-use evolution AKITO Y. KAWAHARA1, DAVID PLOTKIN1,2, ISSEI OHSHIMA3, CARLOS LOPEZ-VAAMONDE4,5, PETER R. HOULIHAN1,6, JESSE W. BREINHOLT1, ATSUSHI KAWAKITA7, LEI XIAO1,JEROMEC. REGIER8,9, DONALD R. DAVIS10, TOSIO KUMATA11, JAE-CHEON SOHN9,10,12, JURATE DE PRINS13 andCHARLES MITTER9 1Florida Museum of Natural History, University of Florida, Gainesville, FL, U.S.A., 2Department of Entomology and Nematology, University of Florida, Gainesville, FL, U.S.A., 3Kyoto Prefectural University, Kyoto, Japan, 4INRA, UR0633 Zoologie Forestière, Orléans, France, 5IRBI, UMR 7261, CNRS/Université François-Rabelais de Tours, Tours, France, 6Department of Biology, University of Florida, Gainesville, FL, U.S.A., 7Center for Ecological Research, Kyoto University, Kyoto, Japan, 8Institute for Bioscience and Biotechnology Research, University of Maryland, College Park, MD, U.S.A., 9Department of Entomology, University of Maryland, College Park, MD, U.S.A., 10Department of Entomology, National Museum of Natural History, Smithsonian Institution, Washington, DC, U.S.A., 11Hokkaido University Museum, Sapporo, Japan, 12Department of Environmental Education, Mokpo National University, Muan, South Korea and 13Department of Entomology, Royal Belgian Institute of Natural Sciences, Brussels, Belgium Abstract. Gracillariidae are one of the most diverse families of internally feeding insects, and many species are economically important. Study of this family has been hampered by lack of a robust and comprehensive phylogeny. In the present paper, we sequenced up to 22 genes in 96 gracillariid species, representing all previously rec- ognized subfamilies and genus groups, plus 20 outgroups representing other families and superfamilies.