Montane Meadows of the Sierra Nevada
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mangrove Swamp (Caroni Wetland, Trinidad)
FIGURE 1.3 Swamps. (a) Floodplain swamp (Ottawa River, Canada). (b) Mangrove swamp (Caroni wetland, Trinidad). FIGURE 1.4 Marshes. (a) Riverine marsh (Ottawa River, Canada; courtesy B. Shipley). (b) Salt marsh (Petpeswick Inlet, Canada). FIGURE 1.5 Bogs. (a) Lowland continental bog (Algonquin Park, Canada). (b) Upland coastal bog (Cape Breton Island, Canada). FIGURE 1.6 Fens. (a) Patterned fen (northern Canada; courtesy C. Rubec). (b) Shoreline fen (Lake Ontario, Canada). FIGURE 1.7 Wet meadows. (a) Sand spit (Long Point, Lake Ontario, Canada; courtesy A. Reznicek). (b) Gravel lakeshore (Tusket River, Canada; courtesy A. Payne). FIGURE 1.8 Shallow water. (a) Bay (Lake Erie, Canada; courtesy A. Reznicek). (b) Pond (interdunal pools on Sable Island, Canada). FIGURE 2.1 Flooding is a natural process in landscapes. When humans build cities in or adjacent to wetlands, flooding can be expected. This example shows Cedar Rapids in the United States in 2008 (The Gazette), but incidences of flood damage to cities go far back in history to early cities such as Nineveh mentioned in The Epic of Gilgamesh (Sanders 1972). FIGURE 2.5 Many wetland organisms are dependent upon annual flood pulses. Animals discussed here include (a) white ibis (U.S. Fish and Wildlife Service), (b) Mississippi gopher frog (courtesy M. Redmer), (c) dragonfly (courtesy C. Rubec), and (d) tambaqui (courtesy M. Goulding). Plants discussed here include (e) furbish lousewort (bottom left; U.S. Fish and Wildlife Service) and ( f ) Plymouth gentian. -N- FIGURE 2.10 Spring floods produce the extensive bottomland forests that accompany many large rivers, such as those of the southeastern United States of America. -

Wetland Valuation Volume I Wetland Ecosystem Services and Their
Wetland Health and Importance Research Programme Wetland Valuation Volume I 8 Wetland ecosystem services and their valuation: a review of current understanding and practice TT 440/09 Wetland Health and Importance Research Programme – 8 Health and Importance Programme Research TTWetland 440/09 Authors: J Turpie, K Lannas, N Scovronick & A Louw Series Editor: H Malan TT 440/09 WETLAND HEALTH AND IMPORTANCE RESEARCH PROGRAMME 8 WETLAND VALUATION. VOL I WETLAND ECOSYSTEM SERVICES AND THEIR VALUATION: A REVIEW OF CURRENT UNDERSTANDING AND PRACTICE Report to the Water Research Commission by Authors: J Turpie1, K Lannas2, N Scovronick1 and A Louw1 Series Editor: H Malan2 1 Anchor Environmental Consultants in association with the Percy FitzPatrick Institute, University of Cape Town 2 Freshwater Research Unit, University of Cape Town WRC Report No. TT 440/09 March 2010 OBTAINABLE FROM Water Research Commission Private Bag X03 Gezina, 0031 The publication of this report emanates from a project entitled Wetland Health and Importance Research Programme (WRC Project no. K5/1584) DISCLAIMER This report has been reviewed by the Water Research Commission (WRC) and approved for publication. Approval does not signify that the contents necessarily reflect the views and policies of the WRC, nor does mention of trade names or commercial products constitute endorsement or recommendation for use. ISBN 978-1-77005-932-0 Set No. 978-1-77005-936-8 Printed in the Republic of South Africa Front Cover: View from bird hide looking upstream, Nylsvley Nature Reserve, Limpopo Province, South Africa Photograph: J Turpie i PREFACE This report is one of the outputs of the Wetland Health and Importance (WHI) research programme which was funded by the Water Research Commission. -

Exploring Our Wonderful Wetlands Publication
Exploring Our Wonderful Wetlands Student Publication Grades 4–7 Dear Wetland Students: Are you ready to explore our wonderful wetlands? We hope so! To help you learn about several types of wetlands in our area, we are taking you on a series of explorations. As you move through the publication, be sure to test your wetland wit and write about wetlands before moving on to the next exploration. By exploring our wonderful wetlands, we hope that you will appreciate where you live and encourage others to help protect our precious natural resources. Let’s begin our exploration now! Southwest Florida Water Management District Exploring Our Wonderful Wetlands Exploration 1 Wading Into Our Wetlands ................................................Page 3 Exploration 2 Searching Our Saltwater Wetlands .................................Page 5 Exploration 3 Finding Out About Our Freshwater Wetlands .............Page 7 Exploration 4 Discovering What Wetlands Do .................................... Page 10 Exploration 5 Becoming Protectors of Our Wetlands ........................Page 14 Wetlands Activities .............................................................Page 17 Websites ................................................................................Page 20 Visit the Southwest Florida Water Management District’s website at WaterMatters.org. Exploration 1 Wading Into Our Wetlands What exactly is a wetland? The scientific and legal definitions of wetlands differ. In 1984, when the Florida Legislature passed a Wetlands Protection Act, they decided to use a plant list containing plants usually found in wetlands. We are very fortunate to have a lot of wetlands in Florida. In fact, Florida has the third largest wetland acreage in the United States. The term wetlands includes a wide variety of aquatic habitats. Wetland ecosystems include swamps, marshes, wet meadows, bogs and fens. Essentially, wetlands are transitional areas between dry uplands and aquatic systems such as lakes, rivers or oceans. -

Laurentian-Acadian Wet Meadow-Shrub Swamp
Laurentian-Acadian Wet Meadow-Shrub Swamp Macrogroup: Wet Meadow / Shrub Marsh yourStateNatural Heritage Ecologist for more information about this habitat. This is modeledmap a distributiononbased current and is data nota substitute for field inventory. based Contact © Maine Natural Areas Program Description: A shrub-dominated swamp or wet meadow on mineral soils characteristic of the glaciated Northeast and scattered areas southward. Examples often occur in association with lakes and ponds or streams, and can be small and solitary pockets or, more often, part of a larger wetland complex. The habitat can have a patchwork of shrub and herb dominance. Typical species include willow, red-osier dogwood, alder, buttonbush, meadowsweet, bluejoint grass, tall sedges, and rushes. Trees are generally absent or thinly scattered. State Distribution: CT, DE, MA, MD, ME, NH, NJ, NY, PA, RI, VA, VT, WV Total Habitat Acreage: 990,077 Ecological Setting and Natural Processes: Percent Conserved: 25.5% Shrub swamps and wet meadows are associated with lakes State State GAP 1&2 GAP 3 Unsecured and ponds and along headwater and larger streams where State Habitat % Acreage (acres) (acres) (acres) the water level does not fluctuate greatly. They are ME 30% 297,075 11,928 39,478 245,668 commonly flooded for part of the growing season but NY 30% 293,979 59,329 38,332 196,318 generally do not have standing water throughout the season. This is a dynamic system that may return to marsh in beaver- MA 8% 76,718 4,358 17,980 54,380 impounded areas or succeed to wooded swamp with NJ 7% 68,351 16,148 9,221 42,983 sediment accumulation or water subsidence. -

1 LAHONTAN MOUNTAIN SUCKER Catostomus Lahontan (Rutter
LAHONTAN MOUNTAIN SUCKER Catostomus lahontan (Rutter) Status: Moderate Concern. The Lahontan mountain sucker does not appear to be at risk of extinction in California in the near future; however, many populations are declining and their range is fragmented. Description: Mountain suckers are small (adults 12-20 cm TL), with subterminal mouths and full lips that are covered by many large papillae (Moyle 2002). Their lips are protrusible, have deep grooves where the upper and lower lips meet, and a cleft on the middle of the lower lip. The lower lip has two semicircular smooth areas along the inner margin next to a conspicuous cartilaginous plate that is used for scraping. The front of the upper lip is smooth. They have 75-92 scales along the lateral line and 23-37 gill rakers on the first gill arch. Fin rays typically number 10 (range 8-13) and nine for the dorsal and pelvic fins, respectively. An axillary process is easily visible at the base of the pelvic fins. Internally, their intestine is long (up to six times TL), and the lining of the abdominal cavity (peritoneum) is black. Their coloration is brown to olive green on the dorsal and lateral surfaces, white to yellow on their bellies, and dark brown in blotches in a lateral row or line. Mature males have two lateral bands, one red-orange on top of another that is black-green. Spawning males have tubercles covering their bodies and fins, with the exception of the dorsal fin. Tubercles on the enlarged anal fin become especially prominent. Spawning females also have tubercles but only on the top and sides of their heads and bodies. -

Riparian Systems
Riparian Systems January 2007 Fish and Wildlife Habitat Management Leaflet Number 45 Introduction Riparian areas are transitional zones between terres- trial and aquatic systems exhibiting characteristics of both systems. They perform vital ecological functions linking terrestrial and aquatic systems within water- sheds. These functions include protecting aquatic eco- systems by removing sediments from surface runoff, decreasing flooding, maintaining appropriate water conditions for aquatic life, and providing organic ma- terial vital for productivity and structure of aquatic ecosystems. They also provide excellent wildlife hab- itat, offering not only a water source, but food and shelter, as well. NRCS Soils in riparian areas differ from soils in upland areas because they are formed from sediments with differ- ent textures and subjected to fluctuating water levels and degrees of wetness. These sediments are rich in nutrients and organic matter which allow the soils to retain large amounts of moisture, affecting the growth and diversity of the plant communities. Riparian areas typically are vegetated with lush growths of grasses, forbs, shrubs, and trees that are tolerant of periodic flooding. In some regions (Great Plains), however, trees may not be part of the his- toric riparian community. Areas with saline soils or U.S. Fish & Wildlife Service heavy, nearly-anaerobic soils (wet meadow environ- ments and high elevations) also are dominated by her- baceous vegetation. In intermittent waterways, the ri- parian area may be confined to the stream channel. Threats to riparian areas have come from many sourc- es. Riparian forests and bottomlands are fertile and valued farmland and rangeland, as well as prime wa- ter-front property desired by developers. -

Wet Meadow Wetland: Wet Meadow
Wet Meadow Wetland: Wet Meadow A wet meadow is an open wetland habitat with soils that are saturated for part or all of the growing season. They have predominantly herbaceous (non-woody) plants that may include any combination of grasses, sedges, rushes, ferns, and forbs. The vegetation may be low and sparse, or tall and dense, depending on a variety of environmental factors. Although standing water may be present at times— especially in winter or spring or after rainstorms—it does not persist for long, and many wet meadows have little if any standing water through much of the growing season. Like other kinds of meadows, many wet meadows are on active or inactive farmland. How to Recognize It Confirming that this scruffy Claverack pasture was wet meadow would require taking a closer look at the As the name implies, the key characteristics are both the plants. wet soils and open, meadow-like conditions. However, during drier parts of the year, the ground underfoot can crunch rather than squelch. Wet meadows can also be as inconspicuous as a small, short patch of bristly sedges Wet Meadow Hand Hollow PCA in the middle of a July pasture or as flowery as a late- Examples summer purple burst of chest-high Purple Loosestrife, Joe-Pye Weed and Blue Vervain. Schor PCA Borden Pond PCA Because of the periodic soggy ground, wet meadows are often not cut or grazed as frequently and closely as drier Mud Creek ELC Beebe Hill fields. Thus, scruffy meadows occurring in local low State Forest spots on the landscape are likely candidates as are ‘roughs’ around landscaped ponds, where lawn mowers Keep Preserve have been deterred by soft ground. -

Guide for Constructed Wetlands
A Maintenance Guide for Constructed of the Southern WetlandsCoastal Plain Cover The constructed wetland featured on the cover was designed and photographed by Verdant Enterprises. Photographs Photographs in this books were taken by Christa Frangiamore Hayes, unless otherwise noted. Illustrations Illustrations for this publication were taken from the works of early naturalists and illustrators exploring the fauna and flora of the Southeast. Legacy of Abundance We have in our keeping a legacy of abundant, beautiful, and healthy natural communities. Human habitat often closely borders important natural wetland communities, and the way that we use these spaces—whether it’s a back yard or a public park—can reflect, celebrate, and protect nearby natural landscapes. Plant your garden to support this biologically rich region, and let native plant communities and ecologies inspire your landscape. A Maintenance Guide for Constructed of the Southern WetlandsCoastal Plain Thomas Angell Christa F. Hayes Katherine Perry 2015 Acknowledgments Our thanks to the following for their support of this wetland management guide: National Oceanic and Atmospheric Administration (grant award #NA14NOS4190117), Georgia Department of Natural Resources (Coastal Resources and Wildlife Divisions), Coastal WildScapes, City of Midway, and Verdant Enterprises. Additionally, we would like to acknowledge The Nature Conservancy & The Orianne Society for their partnership. The statements, findings, conclusions, and recommendations are those of the author(s) and do not necessarily reflect the views of DNR, OCRM or NOAA. We would also like to thank the following professionals for their thoughtful input and review of this manual: Terrell Chipp Scott Coleman Sonny Emmert Tom Havens Jessica Higgins John Jensen Christi Lambert Eamonn Leonard Jan McKinnon Tara Merrill Jim Renner Dirk Stevenson Theresa Thom Lucy Thomas Jacob Thompson Mayor Clemontine F. -

Wet Meadow and Riparian Restoration and Resilience-Building Project 1
Gunnison Basin Wet Meadow and Riparian Restoration and Resilience-Building Project 1. CRITICAL WILDLIFE HABITAT Wet meadows and riparian areas occupy a small proportion of the sagebrush ecosystem in the Gunnison Basin, yet provide critically important habitat for many species. These mesic areas, in an otherwise semi-arid climate, are vital to the life cycle of the federally threatened Gunnison sage-grouse, providing important brood-rearing habitat for the grouse to raise their young chicks. These meadows also provide important food and cover for insects, pollinators, neo-tropical migratory birds, mule deer, elk, and forage for domestic livestock. Wet meadows and riparian areas also serve as natural sponges to hold water in the soil, slowly releasing it after runoff events, ensuring continued base flows and maintenance of water tables throughout the growing season. Holding water in these systems later into the summer season and during droughts benefits plants, wildlife, and ranchers. 2. THREATS TO RESOURCES Many riparian areas and wet meadows in the Gunnison Basin have been degraded by erosion and past land uses, resulting in incised gullies and lowered water tables. Intense episodic droughts, such as the one in 2002, have exacerbated these problems. As our temperatures continue to rise, droughts and intense runoff events that increase erosion are likely to become more frequent. If these already uncommon habitats degrade further, the many plants and animals that depend on them will suffer, including the Gunnison sage-grouse. Since only about 5,000 of these birds remain in the world, they cannot withstand more loss of this important habitat. Degraded riparian areas and wet meadows also pose challenges to ranchers, who are already experiencing other pressures. -
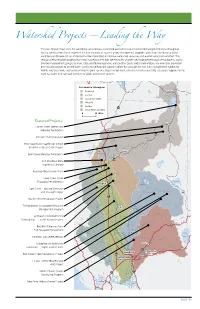
Watershed Projects—Leading The
Watershed Projects—Leading the Way The past 10 years have seen the completion of numerous watershed assessments and watershed management plans throughout the Sacramento River Basin. However, the true measure of success of any management program comes from the ability to affect conditions on the ground, i.e., implement actions to protect or improve watershed resources and overall watershed condition. This section briefly describes projects from each subregion area that are examples of watershed improvement work being done by locally directed management groups; by local, state, and federal agencies; and by other public and private entities. The examples presented here include projects to benefit water quality, streamflow and aquatic habitat, fish passage, fire and fuels management, habitat for wildlife and waterfowl, eradication of invasive plant species, flood management, and watershed stewardship education. Support for this work has come from a broad spectrum of public and private sources. Sacramento Subregions Northeast Lakeview Eastside OREGON Sacramento Valley CALIFORNIA Westside 5 Goose Feather 97 Lake Yuba, American & Bear 0 20 Miles Featured Projects: Alturas Lassen Creek Stream and Mt Shasta r Meadow Restoration e v i R 299 395 t i Pit River Channel Erosion P r e er iv iv R R o t RCD Cooperative Sagebrush Steppe n e m a r d c u Iniatitive — Butte Creek Project a o r S l 101 C e v c i R Lake M Burney Shasta Bear Creek Meadow Restoration Pit 299 CA NEVA Iron Mountain Mine LIFORNIA Eagle Superfund Cleanup Lake Redding DA Redding Allied Stream Team d Cr. Cottonwoo Lower Clear Creek Floodway Rehabilitation Honey Lake Red Bluff Lake Almanor Cow Creek— Bassett Diversion Fish Passage Project 395 r. -

The Historical Range of Beaver in the Sierra Nevada: a Review of the Evidence
Spring 2012 65 California Fish and Game 98(2):65-80; 2012 The historical range of beaver in the Sierra Nevada: a review of the evidence RICHARD B. LANMAN*, HEIDI PERRYMAN, BROCK DOLMAN, AND CHARLES D. JAMES Institute for Historical Ecology, 556 Van Buren Street, Los Altos, CA 94022, USA (RBL) Worth a Dam, 3704 Mt. Diablo Road, Lafayette, CA 94549, USA (HP) OAEC WATER Institute, 15290 Coleman Valley Road, Occidental, CA 95465, USA (BD) Bureau of Indian Affairs, Northwest Region, Branch of Environmental and Cultural Resources Management, Portland, OR 97232, USA (CDJ) *Correspondent: [email protected] The North American beaver (Castor canadensis) has not been considered native to the mid- or high-elevations of the western Sierra Nevada or along its eastern slope, although this mountain range is adjacent to the mammal’s historical range in the Pit, Sacramento and San Joaquin rivers and their tributaries. Current California and Nevada beaver management policies appear to rest on assertions that date from the first half of the twentieth century. This review challenges those long-held assumptions. Novel physical evidence of ancient beaver dams in the north central Sierra (James and Lanman 2012) is here supported by a contemporary and expanded re-evaluation of historical records of occurrence by additional reliable observers, as well as new sources of indirect evidence including newspaper accounts, geographical place names, Native American ethnographic information, and assessments of habitat suitability. Understanding that beaver are native to the Sierra Nevada is important to contemporary management of rapidly expanding beaver populations. These populations were established by translocation, and have been shown to have beneficial effects on fish abundance and diversity in the Sierra Nevada, to stabilize stream incision in montane meadows, and to reduce discharge of nitrogen, phosphorus and sediment loads into fragile water bodies such as Lake Tahoe. -

Plumas National Forest EA EIS CE
Schedule of Proposed Action (SOPA) 04/01/2008 to 06/30/2008 Plumas National Forest This report contains the best available information at the time of publication. Questions may be directed to the Project Contact. Expected Project Name Project Purpose Planning Status Decision Implementation Project Contact Projects Occurring Nationwide Aerial Application of Fire - Fuels management Completed Actual: 02/18/2008 10/2007 Christopher Wehrli Retardant 10/11/2007 202-205-1332 EA [email protected] Description: The Forest Service proposes to continue the aerial application of fire retardant to fight fires on National Forest System lands. An environmental analysis will be conducted to prepare an Environmental Assessment on the proposed action. Web Link: http://www.fs.fed.us/fire/retardant/index.html Location: UNIT - All Districts-level Units. STATE - All States. COUNTY - All Counties. Nation Wide. National Forest System Land - Regulations, Directives, In Progress: Expected:03/2008 03/2008 Gina Owens Management Planning - Orders FEIS NOA in Federal Register 202-205-1187 Proposed Rule 02/15/2008 [email protected] EIS Description: The Agency proposes to publish a rule at 36 CFR part 219 to finish rulemaking on the land management planning rule issued on January 5, 2005 (2005 rule). The 2005 rule guides development, revision, and amendment of land management plans. Web Link: http://www.fs.fed.us/emc/nfma/2008_planning_rule.html Location: UNIT - All Districts-level Units. STATE - All States. COUNTY - All Counties. LEGAL - All units of the National Forest System. Agency-wide. Plumas National Forest, Forestwide (excluding Projects occurring in more than one Forest) R5 - Pacific Southwest Region Backcountry Discovery Trail - Recreation management Completed Actual: 01/24/2008 08/2008 Peggy Gustafson CE 530-283-7620 [email protected] Description: Designation of Backcountry Discovery Trail (BCDT) on existing roads within the Plumas National Forest to tie together statewide motorized trail Location: UNIT - Plumas National Forest All Units.