The Modern Man: a Revision of His Definition and a New Estimation of His Emergence Date
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Dates of the Discovery of the First Peking Man Fossil Teeth
The Dates of the Discovery of the First Peking Man Fossil Teeth Qian WANG,LiSUN, and Jan Ove R. EBBESTAD ABSTRACT Four teeth of Peking Man from Zhoukoudian, excavated by Otto Zdansky in 1921 and 1923 and currently housed in the Museum of Evolution at Uppsala University, are among the most treasured finds in palaeoanthropology, not only because of their scientific value but also for their important historical and cultural significance. It is generally acknowledged that the first fossil evidence of Peking Man was two teeth unearthed by Zdansky during his excavations at Zhoukoudian in 1921 and 1923. However, the exact dates and details of their collection and identification have been documented inconsistently in the literature. We reexamine this matter and find that, due to incompleteness and ambiguity of early documentation of the discovery of the first Peking Man teeth, the facts surrounding their collection and identification remain uncertain. Had Zdansky documented and revealed his findings on the earliest occasion, the early history of Zhoukoudian and discoveries of first Peking Man fossils would have been more precisely known and the development of the field of palaeoanthropology in early twentieth century China would have been different. KEYWORDS: Peking Man, Zhoukoudian, tooth, Uppsala University. INTRODUCTION FOUR FOSSIL TEETH IDENTIFIED AS COMING FROM PEKING MAN were excavated by palaeontologist Otto Zdansky in 1921 and 1923 from Zhoukoudian deposits. They have been housed in the Museum of Evolution at Uppsala University in Sweden ever since. These four teeth are among the most treasured finds in palaeoanthropology, not only because of their scientific value but also for their historical and cultural significance. -

Language Evolution to Revolution
Research Ideas and Outcomes 5: e38546 doi: 10.3897/rio.5.e38546 Research Article Language evolution to revolution: the leap from rich-vocabulary non-recursive communication system to recursive language 70,000 years ago was associated with acquisition of a novel component of imagination, called Prefrontal Synthesis, enabled by a mutation that slowed down the prefrontal cortex maturation simultaneously in two or more children – the Romulus and Remus hypothesis Andrey Vyshedskiy ‡ ‡ Boston University, Boston, United States of America Corresponding author: Andrey Vyshedskiy ([email protected]) Reviewable v1 Received: 25 Jul 2019 | Published: 29 Jul 2019 Citation: Vyshedskiy A (2019) Language evolution to revolution: the leap from rich-vocabulary non-recursive communication system to recursive language 70,000 years ago was associated with acquisition of a novel component of imagination, called Prefrontal Synthesis, enabled by a mutation that slowed down the prefrontal cortex maturation simultaneously in two or more children – the Romulus and Remus hypothesis. Research Ideas and Outcomes 5: e38546. https://doi.org/10.3897/rio.5.e38546 Abstract There is an overwhelming archeological and genetic evidence that modern speech apparatus was acquired by hominins by 600,000 years ago. On the other hand, artifacts signifying modern imagination, such as (1) composite figurative arts, (2) bone needles with an eye, (3) construction of dwellings, and (4) elaborate burials arose not earlier than © Vyshedskiy A. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. -

Verhaegen M. the Aquatic Ape Evolves
HUMAN EVOLUTION Vol. 28 n.3-4 (237-266) - 2013 Verhaegen M. The Aquatic Ape Evolves: Common Miscon- Study Center for Anthropology, ceptions and Unproven Assumptions About Mechelbaan 338, 2580 Putte, the So-Called Aquatic Ape Hypothesis Belgium E-mail: [email protected] While some paleo-anthropologists remain skeptical, data from diverse biological and anthropological disciplines leave little doubt that human ancestors were at some point in our past semi- aquatic: wading, swimming and/or diving in shallow waters in search of waterside or aquatic foods. However, the exact sce- nario — how, where and when these semi-aquatic adaptations happened, how profound they were, and how they fit into the KEY WORDS: human evolution, hominid fossil record — is still disputed, even among anthro- Littoral theory, Aquarboreal pologists who assume some semi-aquatic adaptations. theory, aquatic ape, AAT, Here, I argue that the most intense phase(s) of semi-aquatic Archaic Homo, Homo erectus, adaptation in human ancestry occurred when populations be- Neanderthal, bipedalism, speech longing to the genus Homo adapted to slow and shallow littoral origins, Alister Hardy, Elaine diving for sessile foods such as shellfish during part(s) of the Morgan, comparative biology, Pleistocene epoch (Ice Ages), possibly along African or South- pachyosteosclerosis. Asian coasts. Introduction The term aquatic ape gives an incorrect impression of our semi-aquatic ancestors. Better terms are in my opinion the coastal dispersal model (Munro, 2010) or the littoral theory of human evolution, but although littoral seems to be a more appropriate biologi- cal term here than aquatic, throughout this paper I will use the well-known and common- ly used term AAH as shorthand for all sorts of waterside and semi-aquatic hypotheses. -

Java Man and the Politics of Natural History an Object Biography
Bijdragen tot de Taal-, Land- en Volkenkunde 177 (2021) 290–311 bki brill.com/bki Java Man and the Politics of Natural History An Object Biography Caroline Drieënhuizen | orcid: 0000-0002-5719-2328 Open University of the Netherlands, Heerlen, the Netherlands [email protected] Fenneke Sysling | orcid: 0000-0001-8097-1568 Leiden University, Leiden, the Netherlands [email protected] Abstract Natural history museums have long escaped postcolonial or decolonial scrutiny; their specimens were and are usually presented as part of the natural world, containing only biological or geological information. However, their collections, like those of other museums, are rooted in colonial practices and thinking. In this article, we sketch a polit- ical and decolonial biography of ‘Java Man’, the fossilized remains of a Homo erectus specimen, housed in Naturalis, the Natural History Museum, in the Netherlands. We describe the context of Dutch colonialism and the role of indigenous knowledge and activity in the discovery of Java Man.We also follow Java Man to the Netherlands, where it became a contested specimen and part of a discussion about repatriation. This art- icle argues that the fossils of Java Man and their meanings are products of ‘creolized’ knowledge systems produced by Empire and sites of competing national and discip- linary histories and identities. Keywords natural history museums – Java Man – repatriation – object biography © caroline drieënhuizen and fenneke sysling, 2021 | doi:10.1163/22134379-bja10012 This is an open access article distributed under the terms of the cc by-ncDownloaded4.0 license. from Brill.com09/29/2021 05:34:48PM via free access java man and the politics of natural history 291 1 Introduction Natural history museums have long escaped postcolonial or decolonial scrutiny and are only now somewhat hesitantly joining the conversation about the role of non-Western objects in Western museums that has hitherto focused on eth- nological and art museums. -

The Evolution of Language: Towards Gestural Hypotheses, 208 S
This is a submitted manuscript version. The publisher should be contacted for permission to re-use or reprint the material in any form. Final published version, copyright Peter Lang Publishing: https://www.peterlang.com/view/title/62447 The Evolution of Language: Towards Gestural Hypotheses Przemysław Żywiczyński and Sławomir Wacewicz A publication of the Center for Language Evolution Studies (CLES) NCU Toruń Contents Introduction to the Translation 4 Introduction 5 Chapter 1 7 The Beginnings of Language and Language Origins 7 1.1. Religious beginnings 7 1.1.1. On the divinity of language, the forbidden experiment, and the Adamic language 8 1.1.2. Language as the object of investigation 9 1.1.3. Reflections on language in Indian philosophy 13 1.1.4. Summary 15 1.2. Glottogenetic thought: A naturalistic concept of language emergence 16 1.2.1. How to recover from the state of nature? 16 1.2.2. Darwin: The beginnings of the science on the evolutionary origin of language 34 1.3. Conclusion 41 Chapter 2 41 Evolution, Evolutionism, Evolutionary Thinking 41 2.1. Evolution and Natural Selection 43 2.1.1. Adaptation 47 2.1.2. Gene’s Eye View and Inclusive Fitness 48 2.2. Universal Darwinism and Cultural Evolution 50 2.3. Evolutionary Psychology 52 1 This is a submitted manuscript version. The publisher should be contacted for permission to re-use or reprint the material in any form. Final published version, copyright Peter Lang Publishing: https://www.peterlang.com/view/title/62447 2.4. Popular Reception and the Sins of Evolutionism 55 2.5. -

Homo Erectus Erectus: the Search for His Artifacts Bartstra, G J
University of Groningen Homo erectus erectus: The Search for His Artifacts Bartstra, G J Published in: Current Anthropology IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 1982 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): Bartstra, G. J. (1982). Homo erectus erectus: The Search for His Artifacts. Current Anthropology, 23(3), 318-320. Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 25-09-2021 Homo Erectus Erectus: The Search for His Artifacts Author(s): Gert-Jan Bartstra Source: Current Anthropology, Vol. 23, No. 3 (Jun., 1982), pp. 318-320 Published by: The University of Chicago Press on behalf of Wenner-Gren Foundation for Anthropological Research Stable URL: https://www.jstor.org/stable/2742319 Accessed: 29-10-2018 11:17 UTC JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. -

Outline 22: Hominid Fossil Record
Outline 22: Hominid Fossil Record Human ancestors A.=Australopithicus Assumed direct lineage to modern humans Babcock textbook Collecting hominid fossils in East Africa Using Stratigraphy and Radiometric Dating of ash beds to date hominid fossils A Hominid Jawbones from Ethiopia Sahelanthropus tchadensis, 6.5 MY old Sahelanthropus tchadensis, 6.5 MY old Gorilla – female on left, male on right; note the sexual dimorphism Orrorin tugenensis – the sum total of specimens Ardipithecus ramidus, 2009 Australopithecus anamensis The Australopithecines • Ardipithecus: oldest definite bipedal ancestor, over 4 M.Y. old • Australopithecus: the gracile australopithecines • Paranthropus: the robust australopithecines Fossil Species • Ardipithecus ramidus: 4.4 MY, teeth, jaws and bone fragments suggest it is a hominid, not a pongid. • Australopithecus afarensis: 3.8-3.0 MY based on good fossils. Clearly a bipedal animal based on bones and fossil footprints. Bipedal Footprints of Australopithecus in Lithified Volcanic Ash 4 MY old from Tanzania. Found by Mary Leakey. Australopithecus couple making footprints in volcanic ash Jawbones of Australopithecus afarensis Fossil remains of “Lucy”, the most complete specimen of Australopithecus afarensis. An adult skull, A. afarensis Pelvis of Australopithecus afarensis Reconstruction of Australopithecus afarensis Face-to-face with A. afarensis A troop of A. afarensis feeding on tubers in the forest. A. afarensis family unit crossing the savannah. Fossil Species • Australopithecus africanus: 2.8-2.5 MY • Robust australopithecines: Paranthropus aethiopicus: 2.6-2.2 MY P. robustus: 2.0-1.2 MY P. boisei: 2.6-1.0 MY The Taung Child, A. africanus Skull of Paranthropus boisei Paranthropus aethiopicus Paranthropus aethiopicus Justus Erus found the fossil Kenyanthropus platyops, 3.5 MY old from Kenya Kenyanthropus rudolfensis or Homo rudolfensis? Fossil Species • Homo habilis: 2.5-1.6 MY • H. -

Development Team
Paper No. : 01 Physical/Biological Anthropology Module : 18 Homo erectus Development Team Prof. Anup Kumar Kapoor Principal Investigator Department of Anthropology, University of Delhi Paper Coordinator Prof. Subho Roy Department of Anthropology ,University of Calcutta Dr. Arpita Mandal (Nandi) Content Writer Department of Anthropology, Narsinha Dutt College, Howrah Content Reviewer Prof. Barun Mukhopadhyay Indian Statistical Institute, Kolkata 1 Physical/ Biological Anthropology Anthropology Homo erectus Description Of Module Subject Name Anthropology Paper Name 01 Physical/ Biological Anthropology Module Name/Title Homo erectus Module Id 18 2 Physical/ Biological Anthropology Anthropology Homo erectus Contents of this unit About Homo erectus Variations and their Distribution Time Period Anatomical Features Cultural Remains Other Aspects of Culture Dispersal of Homo erectus Ascribed the Status of Being the First Behavioral Inferences Phylogenetic / Evolutionary Relationships Theories of Change Learning Objectives Who is Homo erectus? Distributions Important Position in Human Evolution 3 Physical/ Biological Anthropology Anthropology Homo erectus Homo erectus Source: Photobucket.com Introduction The first fossil was found from Java in 1891 by the Dutch anatomist, Eugene Dubois. The exact locale was Trinil river bed in central java, and its common name assigned is “Java man”. The materials found were: a skull cap, a complete femur, three teeth, and a jaw fragment. This specimen was formerly referred to as “Pithecanthropus erectus” meaning an apeman who could walk straight. The femur found among the other materials showed prominent linea aspera, a criterion for erect posture and bipedalism while the skull cap denoted brain volume as that of present day apes. The dating of this fossil is middle pleistocene about 500,000 years ago. -

Evolution of Homo and Related Hominins
Biology 1B – Evolution Lecture 13 – transitional forms Evolution of Homo and related hominins Subfamily Homininae = monophyletic group consisting of humans, chimpanzees & gorillas and species now extinct within each of these lineages. The fossil record provides evidence of the evolution of the Hominin lineage leading to Homo sapiens. There were two main schools of thought regarding the origin of Homo sapiens: Darwin thought humanity arose from Africa (due to presence of chimpanzees and gorillas), while other prominent scientists thought that humans arose out of Asia (due to presence of orangutans and early fossil Homo). Early applications of the molecular clock (by Sarich and Wilson at UC Berkeley) suggest the split between the lineages leading to living chimpanzees on one branch and humans on the other occurred about 7 million years (mya) ago: the last common ancestor between chimpanzees and humans existed around 7mya. Therefore, humans are not descended from chimpanzees: the chimpanzee lineage has been evolving independently as long as our lineage has. Our common ancestor did not look like either chimpanzees or humans: for example, consider Ardepithecus ramidis from 4mya (discovered by Tim White, and visible in the display outside the Marian Koshland Biosciences Library). It is our ancestor, yet it does not look like us. It is neither a chimpanzee nor a human. The Australopithecines are probably paraphyletic with the Homo line. When looking at the graph below, note that it is not a phylogeny or even a cladogram: there is still much debate on the phylogeny and even the taxonomy of these different species. Biology 1B – Evolution Lecture 13 – transitional forms Figure 1: Plot of different hominins and their temporal location Campbell, Fig 34.40, Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings Key features shared by the lineage leading to humans include bipedalism (walking upright on two limbs) and smaller canines. -

Unit 7 Homo Erectus from Africa, Asia, Europe*
Homo Erectus from Africa, UNIT 7 HOMO ERECTUS FROM AFRICA, Asia, Europe ASIA, EUROPE* Contents 7.0 Introduction 7.1 Distribution of Homo erectus 7.1.1 Homo erectus from Java 7.1.2 Homo erectus from China 7.1.3 Homo erectus from Africa 7.1.4 Homo erectus from Europe 7.2 Morphological Features of Homo erectus 7.3 Phylogenetic Status and Life ways of Homo erectus 7.4 Overview of Life History and Biology of Homo erectus 7.5 Summary 7.6 References 7.7 Answers to Check Your Progress Learning Objectives After reading this unit, you will be able to: know the distribution of Homo Erectus in asia, europe, africa; learn about morphological features of Homo Erectus; and understand life history and Biology of Homo Erectus. 7.0 INTRODUCTION The earliest members of the genus Homo are of particular significance as they mark the evolution of our own species. Homo erectus was a true human as he exhibited all the basic elements of the human bio-cultural adaptive pattern. He also used fire, made stone tools and other implements, used the clean skin of other animals and therefore may be taken to represent the beginning of true man. In 1961, L.S.B Leaky reported parts of the skull and jaw of juvenile Hominid from Bed I at Olduvai Gorge. This material came from a site lower in the deposit which yielded Zinjanthropus and hence it was named as Pre-Zinjanthropus. In 1963, Leaky recovered more fossil material from Bed I and II from Olduvai. In 1972, Richard Leaky, showed casts of fossils from deposits at Lake Turkana to which he assigned a third Hominid model. -

Sociology and Anthropology: the FAMILY TREE of MAN
Cebu Normal University COLLEGE OF NURSING Osmeña Boulevard, Cebu City Sociology and Anthropology: THE FAMILY TREE OF MAN Australopithecus Homo Erectus Javanensis (Java Man) Homo Erectus Pekinensis (Peking Man) Neanderthal Man Andales, Elisha Gine Nuez, Willy Angelie ALjaberi, Khamis Fahad BSN IV The Family Tree of Man Human evolution is the evolutionary process leading up to the appearance of modern humans. While it began with the last common ancestor of all life, the topic usually covers only the evolutionary history of primates, in particular the genus Homo, and the emergence of Homo sapiens as a distinct species of hominids (or "great apes"). The study of human evolution involves many scientific disciplines, including physical anthropology, primatology, archaeology, embryology and genetics. A number of theories exist about the evolution of man, one of which is Charles Darwin’s Theory of Natural Selection. In this theory, Darwin noted that individual members of a species differ from one another physically. For instance, in a given population, some animals may have longer limbs than others. These variations are, of course, dependent upon the demands placed on the organism by its environment. Moreover, it enhances the animal’s chance of producing offsprings that survive. In effect, with each generation, the better adapt members of a population increase in number at the expense of less favored individuals. In the process too, the species as a whole change (Plog, 1980.13). This theory tends to explain not only gradual change within the species but also the appearance of new species. In addition, the theory states that adaptation to different environmental circumstances accounts for great variety of species observable in nature. -
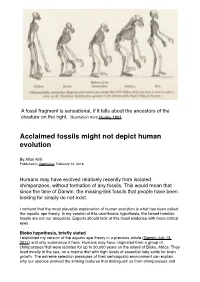
Acclaimed Fossils Might Not Depict Human Evolution
Page 1! A fossil fragment is sensational, if it tells about the ancestors of the creature on the right. Illustration from Huxley 1863 Acclaimed fossils might not depict human evolution By Allan Krill Published in Gemini.no, February 12, 2018 Humans may have evolved relatively recently from isolated chimpanzees, without formation of any fossils. This would mean that since the time of Darwin, the missing-link fossils that people have been looking for simply do not exist. I contend that the most plausible explanation of human evolution is what has been called the aquatic ape theory. In my version of this unorthodox hypothesis, the famed hominin fossils are not our ancestors. Experts should look at this fossil evidence with more critical eyes. Bioko hypothesis, briefly stated I explained my version of the aquatic ape theory in a previous article (Gemini July 13, 2017) and only summarize it here. Humans may have originated from a group of chimpanzees that were isolated for up to 30,000 years on the island of Bioko, Africa. They lived mostly in the sea, on a marine diet with high levels of essential fatty acids for brain growth. The extreme selection pressures of their semiaquatic environment can explain why our species evolved the striking features that distinguish us from chimpanzees and Page 2! other apes, such as our large brain, naked skin, external nose, long legs, long scalp hair, and newborns with weak neck and baby blubber-fat. No fossils would have formed along the coast of Bioko, because it was an erosional geologic environment without deposition of sediments.