O'odham Resource Use in the Colonial Pimería Alta
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
2020 Monterey County Ranch Map Atlas 89 Pages Standard
2D 2E 2F TOM BENGARD RANCH INC. D'ARRIGO BROS. CO. OF CALIFORNIA 14S03E35 WEST HANSEN RANCH 52 14S03E36 RANCH 22 S USDA AG RESEARCH STATION 14S04E31 a HARTNELL RANCH-USDA D'ARRIGO BROS. CO. OF CALIFORNIA 14S04E32 l RANCH 22 PEZZINI BERRY FARMS 14S04E33 i BE BERRY FARMS 14S04E34 n GAMBETTA RANCH HARTNELL RANCH ALLAN W. JOHNSON & SON-ORGANIC a HOME RANCH s A i r TRIANGLE FARMS INC. ROBERT SILVA FARMS (ORGANIC) p HARTNELL RANCH o WILLIAMS/DAVIS/MILLER RANCH 07 o D'ARRIGO BROS. CO. OF CALIFORNIA r ROBERT SILVA FARMS (ORGANIC) RANCH 22 t CHRISTENSEN & GIANNINI LLC. A DAGGETT/HEDBERG SOUTH MORTENSEN RANCH L A B B ALLAN W. JOHNSON & SON A A Z FUENTES FARMS R JOHNSON & SON HOME RANCH LAURITSON RANCH D IN BUCIO FARMS ORGANIC RICKY'S FARMS SAN ANTONIO RANCH 184 ZABALA RD. 15S03E02 SUNLIGHT BERRY FARMS INC. 15S03E01 15S04E06 LAURITSON RANCH MERRILL FARMS LLC. - VEGETABLE 15S04E05 O CHRISTENSEN & GIANNINI LLC. ALLAN W. JOHNSON & SON L 15S04E04 NORTON RANCH D ALISAL RANCH WILSON RANCH 15S04E03 ROBERT SILVA FARMS S T LAURITSON RANCH A ALLAN W. JOHNSON & SON NIXON RANCH G E MERRILL FARMS LLC. - VEGETABLE ALLAN W. JOHNSON & SON-ORGANIC AIRPORT RANCH NIXON ORGANIC RANCH ALLAN W. JOHNSON & SON-ORGANIC NIXON ORGANIC RANCH G & H FARMS LLC. ORGANIC CHRISTENSEN & GIANNINI LLC. ALISAL RANCH CUMMINGS RANCH ALLAN W. JOHNSON & SON G & H FARMS GONZALEZ ORGANIC FARMS NIXON RANCH ALLAN W. JOHNSON & SON BARDIN RANCH ZABALA RANCH NIXON RANCH GONZALEZ ORGANIC FARMS MORESCO FARMS INC. SUN COAST GROWERS G & H FARMS GONZALEZ RANCH ALISAL RANCH HARDEN RANCH 5 ROBERT SILVA FARMS ZABALA ROAD BAY FRESH PRODUCER ZABALA RANCH 10 ROBERT SILVA FARMS HARTNELL RANCH SUN COAST RANCH 11 15S03E11 15S03E12 15S04E07 ROBERT SILVA FARMS GARCIA HOME RANCH 2 15S04E08 L 15S04E09 D'ARRIGO BROS. -

Sample Costs for Beef Cattle, Cow-Calf Production
UNIVERSITY OF CALIFORNIA AGRICULTURE AND NATURAL RESOURCES COOPERATIVE EXTENSION AGRICULTURAL ISSUES CENTER UC DAVIS DEPARTMENT OF AGRICULTURAL AND RESOURCE ECONOMICS SAMPLE COSTS FOR BEEF CATTLE COW – CALF PRODUCTION 300 Head NORTHERN SACRAMENTO VALLEY 2017 Larry C. Forero UC Cooperative Extension Farm Advisor, Shasta County. Roger Ingram UC Cooperative Extension Farm Advisor, Placer and Nevada Counties. Glenn A. Nader UC Cooperative Extension Farm Advisor, Sutter/Yuba/Butte Counties. Donald Stewart Staff Research Associate, UC Agricultural Issues Center and Department of Agricultural and Resource Economics, UC Davis Daniel A. Sumner Director, UC Agricultural Issues Center, Costs and Returns Program, Professor, Department of Agricultural and Resource Economics, UC Davis Beef Cattle Cow-Calf Operation Costs & Returns Study Sacramento Valley-2017 UCCE, UC-AIC, UCDAVIS-ARE 1 UC AGRICULTURE AND NATURAL RESOURCES COOPERATIVE EXTENSION AGRICULTURAL ISSUES CENTER UC DAVIS DEPARTMENT OF AGRICULTURAL AND RESOURCE ECONOMICS SAMPLE COSTS FOR BEEF CATTLE COW-CALF PRODUCTION 300 Head Northern Sacramento Valley – 2017 STUDY CONTENTS INTRODUCTION 2 ASSUMPTIONS 3 Production Operations 3 Table A. Operations Calendar 4 Revenue 5 Table B. Monthly Cattle Inventory 6 Cash Overhead 6 Non-Cash Overhead 7 REFERENCES 9 Table 1. COSTS AND RETURNS FOR BEEF COW-CALF PRODUCTION 10 Table 2. MONTHLY COSTS FOR BEEF COW-CALF PRODUCTION 11 Table 3. RANGING ANALYSIS FOR BEEF COW-CALF PRODUCTION 12 Table 4. EQUIPMENT, INVESTMENT AND BUSINESS OVERHEAD 13 INTRODUCTION The cattle industry in California has undergone dramatic changes in the last few decades. Ranchers have experienced increasing costs of production with a lack of corresponding increase in revenue. Issues such as international competition, and opportunities, new regulatory requirements, changing feed costs, changing consumer demand, economies of scale, and competing land uses all affect the economics of ranching. -

Integrated Pest Management: Current and Future Strategies
Integrated Pest Management: Current and Future Strategies Council for Agricultural Science and Technology, Ames, Iowa, USA Printed in the United States of America Cover design by Lynn Ekblad, Different Angles, Ames, Iowa Graphics and layout by Richard Beachler, Instructional Technology Center, Iowa State University, Ames ISBN 1-887383-23-9 ISSN 0194-4088 06 05 04 03 4 3 2 1 Library of Congress Cataloging–in–Publication Data Integrated Pest Management: Current and Future Strategies. p. cm. -- (Task force report, ISSN 0194-4088 ; no. 140) Includes bibliographical references and index. ISBN 1-887383-23-9 (alk. paper) 1. Pests--Integrated control. I. Council for Agricultural Science and Technology. II. Series: Task force report (Council for Agricultural Science and Technology) ; no. 140. SB950.I4573 2003 632'.9--dc21 2003006389 Task Force Report No. 140 June 2003 Council for Agricultural Science and Technology Ames, Iowa, USA Task Force Members Kenneth R. Barker (Chair), Department of Plant Pathology, North Carolina State University, Raleigh Esther Day, American Farmland Trust, DeKalb, Illinois Timothy J. Gibb, Department of Entomology, Purdue University, West Lafayette, Indiana Maud A. Hinchee, ArborGen, Summerville, South Carolina Nancy C. Hinkle, Department of Entomology, University of Georgia, Athens Barry J. Jacobsen, Department of Plant Sciences and Plant Pathology, Montana State University, Bozeman James Knight, Department of Animal and Range Science, Montana State University, Bozeman Kenneth A. Langeland, Department of Agronomy, University of Florida, Institute of Food and Agricultural Sciences, Gainesville Evan Nebeker, Department of Entomology and Plant Pathology, Mississippi State University, Mississippi State David A. Rosenberger, Plant Pathology Department, Cornell University–Hudson Valley Laboratory, High- land, New York Donald P. -

Assessment of the Energy Potential of Chicken Manure in Poland
energies Article Assessment of the Energy Potential of Chicken Manure in Poland Mariusz Ta´nczuk 1,*, Robert Junga 1, Alicja Kolasa-Wi˛ecek 2 and Patrycja Niemiec 1 1 Faculty of Mechanical Engineering, Opole University of Technology, 45-271 Opole, Poland; [email protected] (R.J.); [email protected] (P.N.) 2 Faculty of Natural Sciences and Technology, Opole University, 45-047 Opole, Poland; [email protected] * Correspondence: [email protected]; Tel.: +48-664-475-355 Received: 2 January 2019; Accepted: 25 March 2019; Published: 1 April 2019 Abstract: Animal waste, including chicken manure, is a category of biomass considered for application in the energy industry. Poland is leading poultry producer in Europe, with a chicken population assessed at over 176 million animals. This paper aims to determine the theoretical and technical energy potential of chicken manure in Poland. The volume of chicken manure was assessed as 4.49 million tons per year considering three particular poultry rearing systems. The physicochemical properties of examined manure specimens indicate considerable conformity with the data reported in the literature. The results of proximate and ultimate analyses confirm a considerable effect of the rearing system on the energy parameters of the manure. The heating value of the chicken manure was calculated for the high moisture material in the condition as received from the farms. The value of annual theoretical energy potential in Poland was found to be equal to around 40.38 PJ. Annual technical potential of chicken biomass determined for four different energy conversion paths occurred significantly smaller then theoretical and has the value from 9.01 PJ to 27.3 PJ. -

Free-Range Environment
The requirements listed in this document pertain to shell egg labels that contain the USDA grademark, only. MANDATORY LABELING REQUIREMENTS • It is strongly recommended that you verify applicable State requirements before submitting labels for Federal approval. • FDA = The Federal Food, Drug, and Cosmetic Act (FDCA); the Fair Packaging and Labeling Act (FPLA); and the regulations promulgated under these acts (21 CFR). • USDA = 7 CFR Part 56; AMS 56 Principal Display Panel (PDP): • The term principal display panel means the part of a label that is most likely to be displayed, presented, shown, or examined under customary conditions of display for retail sale. • The principal display panel shall be large enough to accommodate all the mandatory label information required to be placed on the container with clarity and conspicuousness and without obscuring design, vignettes, or crowding. • The Producer, Distributor, or Carton Manufacturer must declare the PDP or alternate PDP’s prior to issuance of a USDA approval number. Contact this office to further discuss the selection of the PDP. PDP = Top View, only. Back Hinge PDP Front Lip PDP = Top View and Front Lip (Alternate Display Panels) Back Hinge PDP Front Lip • If the Top View and Front Lip are selected as the PDP’s, all required mandatory information must be placed on both panels (Top View and Front Lip). • Overwrapped, Sleeved, or Case containers: All mandatory labeling information must be placed on the top panel or information panel. No mandatory labeling requirements may be placed on the bottom panel. Page 1 08/22/2016 Information Panel: • The information panel as it applies to packaged food means that part of the label immediately contiguous and to the right of the principal display panel (PDP) as observed by an individual facing the principal display panel. -
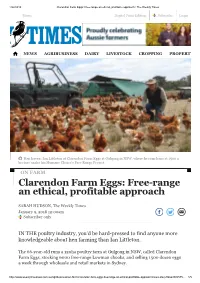
Clarendon Farm Eggs: Free-Range an Ethical, Profitable Approach | the Weekly Times
1/22/2018 Clarendon Farm Eggs: Free-range an ethical, profitable approach | The Weekly Times Menu Digital Print Edition Subscribe Login NEWS AGRIBUSINESS DAIRY LIVESTOCK CROPPING PROPERTY Hen haven: Ian Littleton at Clarendon Farm Eggs at Gulgong in NSW, where he runs hens at 1500 a hectare under his Humane Choice’s Free Range Project. ON FARM Clarendon Farm Eggs: Free-range an ethical, profitable approach SARAH HUDSON, The Weekly Times January 9, 2018 12:00am Subscriber only IN THE poultry industry, you’d be hard-pressed to find anyone more knowledgeable about hen farming than Ian Littleton. The 66-year-old runs a 120ha poultry farm at Gulgong in NSW, called Clarendon Farm Eggs, stocking 6000 free-range Lowman chooks, and selling 1500 dozen eggs a week through wholesale and retail markets in Sydney. http://www.weeklytimesnow.com.au/agribusiness/on-farm/clarendon-farm-eggs-freerange-an-ethical-profitable-approach/news-story/0dae808c575… 1/5 1/22/2018 Clarendon Farm Eggs: Free-range an ethical, profitable approach | The Weekly Times But the property is more of a semi-retirement project, following a six-decade career Menu Digital Print Edition Subscribe Login in the poultry industry, from which his accumulated knowledge has led to best- practice innovation across all farm productions. IAN LITTLETON GULGONG, NSW STNEWSOCKS 6 00AGRIBUSINESS0 free-range Lowm aDAIRYn chooks oLIVESTOCKn 120ha farm CROPPING PROPERTY HAS 14 mobile chook homes SELLS 1500 dozen eggs a week through wholesale and retail markets in Sydney ACCREDITED with Humane Choice’s Free Range Project Ian grew up on a poultry farm near Toowoomba in Queensland, and, after his father passed away and the farm was sold, studied an agricultural degree at Queensland Agricultural College, specialising in poultry production. -

Native Americans in the Cape Fear, by Dr. Jan Davidson
Native Americans in the Cape Fear, By Dr. Jan Davidson Archaeologists believe that Native Americans have lived in what is now the state of North Carolina for more than 13,000 years. These first inhabitants, now called Paleo-Indians by experts, were likely descended from people who came over a then-existing land bridge from Asia.1 Evidence had been found at Town Creek Mound that suggests Indians lived there as early as 11000 B.C.E. Work at another major North Carolinian Paleo-Indian where Indian artifacts have been found in layers of the soil, puts Native Americans on that land before 8000 B.C.E. That site, in North Carolina’s Uwharrie Mountains, near Badin, became an important source of stone that Paleo and Archaic period Indians made into tools such as spears.2 It is harder to know when the first people arrived in the lower Cape Fear. The coastal archaeological record is not as rich as it is in some other regions. In the Paleo-Indian period around 12000 B.C.E., the coast was about 60 miles further out to sea than it is today. So land where Indians might have lived is buried under water. Furthermore, the coastal Cape Fear region’s sandy soils don’t provide a lot of stone for making tools, and stone implements are one of the major ways that archeologists have to trace and track where and when Indians lived before 2000 B.C.E.3 These challenges may help explain why no one has yet found any definitive evidence that Indians were in New Hanover County before 8000 B.C.E.4 We may never know if there were indigenous people here before the Archaic period began in approximately 8000 B.C.E. -

C:\Documents and Settings\David Carlson\Desktop\SHA97
1997 SHA Conference on Historical and Underwater SOCIETY FOR HISTORICAL ARCHAEOLOGY Archaeology Corpus Christi, TX 1997 AWARDS OF MERIT January 8 - 12, 1997 to be presented to PILAR LUNA ERREGUERENA Instituto Nacional de Antropologia e Historia, Mexico Seaports, Ships, and Central Places TEXAS HISTORICAL COMMISSION Abstracts TEXAS ARCHEOLOGICAL SOCIETY 1997 J.C. HARRINGTON MEDAL JAMES DEETZ University of Virginia Hosted by Texas A&M University Institute of Nautical Archaeology Ships of Discovery ABSTRACTS 1997 CONFERENCE STAFF Conference Chair and Program Coordinator............. David L. Carlson Terrestrial Program Chair .......................Shawn Bonath Carlson Underwater Program Chair ............................. Denise Lakey Registration Chair .............................. Frederick M. Hocker Society for Historical Archaeology Local Arrangements Chair............................... Toni Carrell Volunteer Coordinator.................................Becky Jobling Tours Coordinator .....................................Mary Caruso 30th Conference on Historical and Book Room Coordinator .......................... Lawrence E. Babits Underwater Archaeology Employment Coordinator............................... Sarah Mascia Conference Coordinator................................. Tim Riordan Hosted by: Texas A&M University Institute of Nautical Archaeology Ships of Discovery January 8-12, 1997 Omni Bayfront Hotel Corpus Christi, Texas With financial support provided by: Corpus Christi Omni Bayfront Hotel Corpus Christi Area Convention & -

A Time to Print, a Time to Reform
Chapman University Chapman University Digital Commons ESI Working Papers Economic Science Institute 3-2019 A Time to Print, a Time to Reform Lars Boerner Martin-Luther University of Halle-Wittenberg Jared Rubin Chapman University, [email protected] Battista Severgnini Copenhagen Business School Follow this and additional works at: https://digitalcommons.chapman.edu/esi_working_papers Part of the Econometrics Commons, Economic Theory Commons, and the Other Economics Commons Recommended Citation Boerner, L., Rubin, J., & Severgnini, B. (2019). A time to print, a time to reform. ESI Working Paper 19-07. Retrieved from https://digitalcommons.chapman.edu/esi_working_papers/264/ This Article is brought to you for free and open access by the Economic Science Institute at Chapman University Digital Commons. It has been accepted for inclusion in ESI Working Papers by an authorized administrator of Chapman University Digital Commons. For more information, please contact [email protected]. A Time to Print, a Time to Reform Comments Working Paper 19-07 This article is available at Chapman University Digital Commons: https://digitalcommons.chapman.edu/ esi_working_papers/264 A Time to Print, a Time to Reform∗ Lars Boerner† Jared Rubin‡ Battista Severgnini§ Abstract The public mechanical clock and movable type printing press were arguably the most important and complex technologies of the late medieval period. We posit that towns with clocks became upper-tail human capital hubs|clocks required extensive technical know-how and fine mechanical skill. This meant that clock towns were in position to adopt the printing press soon after its invention in 1450, as presses required a simi- lar set of mechanical and technical skills to operate and repair. -

Hitchens Ranch Is One of the Oldest Continuously-Owned and Operated Family Ranches in Routt County
A, C, D, & J. HITCHENS OVERLOOK RANCH U. S. Highway 40, Milner The Hitchens Ranch is one of the oldest continuously-owned and operated family ranches in Routt County. Through the Homestead Act, James Hitchens filed on 160 acres in 1886; later, his son Albert bought surrounding lands, and gradually the ranch grew to 1,980 acres. When James built his two-room log cabin along the stage route between Hayden and Steamboat Springs, he dug nearly three-mile-long trenches to irrigate fields and provide a pond in front of his house with which to water the stage horses that rested in the still-extant barn (see photo). At one time, the original log cabin homestead functioned as a post office. Eventually, James built additions and a second story on the house and sided the residence with clapboards. James later built a two- story home in Milner that provided the area with a larger post office, a pool hall, and a barber shop. Albert, the last of nine children, was born on the ranch in 1892, attended school two miles away, and ranched with his father. Later, he homesteaded adjacent to his parents’ home. Albert had a hernia rupture that prevented him from farming, so he worked at as a barber, assistant postmaster, and a cobbler, but since his first love was ranching, he traveled to Kansas City for a paraffin treatment that was apparently successful: He returned to the ranch, moved all of his homestead buildings onto his parents’ ranch in 1929, divided his two-room log house into a chicken coop and a bunkhouse, expanded the barn, and moved the Milner house by a wagon pulled by 28 horses onto the ranch, and lived on and ranched the property until he died at 93 years of age. -

The Impact of the Columbian Exchange on Native American Populations Katelyn K. Chiolan Introduction This Unit Is Written For
The Impact of the Columbian Exchange on Native American Populations Katelyn K. Chiolan Introduction This unit is written for teachers of history courses, specifically eighth grade United States History I. Through this course, students are expected to learn about the beginnings of American history, starting with Native American culture and ending at the Reconstruction Era. It is crucial for students of this course and grade level to begin using certain historical skills, such as conducting basic independent research. To help teach students the importance and value of independent historical research, this unit focuses on applying the concept of numeracy to social studies by evaluating the credibility and reliability of data found through the students’ investigations. The concept of numeracy is one that has been largely ignored by educators but is useful across all curriculums. When I was first introduced to the term numeracy, I assumed that it was a mathematics idiom and would have virtually no part in my history classroom. However, in a social studies class like United States history, being able to understand where data come from and how to apply them to social constructs is not only helpful but necessary; this skill is at the root of numeracy. Students conducting research projects must be able to approach social issues with the ability to discover the truth behind the myriad of myths and misleading information. We must infuse numeracy education into math and other classes so that we and our students can better understand data in research as well as everyday information. Today’s students live in a time when information is so readily available that it seems almost silly to conduct in-depth research. -

Journal of Arizona History Index
Index to the Journal of Arizona History, W-Z Arizona Historical Society, [email protected] 480-387-5355 NOTE: the index includes two citation formats. The format for Volumes 1-5 is: volume (issue): page number(s) The format for Volumes 6 -54 is: volume: page number(s) W WAACs 36:318; 48:12 photo of 36:401 WAAFs 36:318 Wabash Cattle Company 33:35, 38 Wachholtz, Florence, book edited by, reviewed 18:381-82 Waco Tap 38:136 Wacos (airplanes) 15:334, 380 Waddell, Jack O., book coedited by, reviewed 22:273-74 Waddle, Billy 42:36, 38 Wade, Abner, photo of 28:297 property of 28:284, 286, 288, 294 Wade, Benjamin F. 19:202; 41:267, 274, 279, 280, 281, 282, 284 photo of 41:268 Wade, George A. 22:24 Wade Hampton Mine 23:249-50 Wade, James F. 14:136; 29:170 Wade, John Franklin IV(1)6 Wade, Michael S., book by, listed 24:297 1 Index to the Journal of Arizona History, W-Z Arizona Historical Society, [email protected] 480-387-5355 Wade, Nicholas 31:365, 397 n. 34 Wade, William 43:282 Wadleigh, Atherton B. 20:35-36, 56, 58-59 portrait 20:57 Wadsworth, B. C. 27:443 Wadsworth, Craig 41:328 Wadsworth mine 34:151 Wadsworth mining claim 34:122, 123 Wadsworth, Mr. See Wordsworth, William C. Wadsworth, Nevada 54:389 Wadsworth, William R. V(4)2 Wadsworth, William W. 23:21, 23 Waffle, Edison D. 7:20 n. 26 Wager, Evelyn 39:234 n. 1, 234 n.