The Callerya Group Redefined and Tribe Wisterieae (Fabaceae) Emended Based on Morphology and Data from Nuclear and Chloroplast DNA Sequences
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

LK Audited Per 31 Desember 2016
PT WASKITA KARYA (PERSERO) Tbk PT WASKITA KARYA (PERSERO) Tbk DAN ENTITAS ANAK AND SUBSIDIARIES Daftar Isi Halaman/ Table of Contents Pages Surat Pernyataan Direksi Director’s Statement Letter Laporan Auditor Independen Independent Auditor’s Report Laporan Keuangan Konsolidasian Consolidated Financial Statements Untuk Tahun-tahun yang Berakhir pada For the Years Ended Tanggal 31 Desember 2016 dan 2015 December 31, 2016 and 2015 Laporan Posisi Keuangan Konsolidasian 1 Consolidated Statements of Financial Position Laporan Laba Rugi dan Penghasilan 3 Consolidated Statements of Profit or Loss and Komprehensif Lain Konsolidasian Other Comprehensive Income Laporan Perubahan Ekuitas Konsolidasian 4 Consolidated Statements of Changes in Equity Laporan Arus Kas Konsolidasian 5 Consolidated Statements of Cash Flows Catatan Atas Laporan Keuangan 6 Notes to the Consolidated the Financial Konsolidasian Statements PT WASKITA KARYA (PERSERO) Tbk PT WASKITA KARYA (PERSERO) Tbk DAN ENTITAS ANAK AND SUBSIDIARIES LAPORAN POSISI KEUANGAN CONSOLIDATED STATEMENTS OF KONSOLIDASIAN FINANCIAL POSITION 31 Desember 2016 dan 2015 December 31, 2016 and 2015 (Dalam Rupiah Penuh) (In Full of Rupiah) Catatan/ 2016 2015 *) Notes Rp Rp ASET ASSETS ASET LANCAR CURRENT ASSETS Kas dan Setara Kas 4, 49, 51, 54 10,653,780,768,186 5,511,188,078,778 Cash and Cash Equivalents Investasi Jangka Pendek 5 10,663,933,745 10,663,933,745 Short-Term Investments Piutang Usaha Accounts Receivable Pihak Berelasi 6, 51, 54 1,356,258,910,298 1,174,449,804,260 Related Parties Pihak Ketiga 6 -

Universiti Putra Malaysia Genetic Diversity of Tuba
UNIVERSITI PUTRA MALAYSIA GENETIC DIVERSITY OF TUBA PLANTS, AND TOXICITY OF THEIR ROTENOIDS FORMULATED AS NANO-EMULSION AGAINST Plutella xylostella L. NORHAYU BINTI ASIB FP 2015 42 GENETIC DIVERSITY OF TUBA PLANTS, AND TOXICITY OF THEIR ROTENOIDS FORMULATED AS NANO-EMULSION AGAINST Plutella xylostella L. By NORHAYU BINTI ASIB UPM COPYRIGHT © Thesis presented to the Senate of Universiti Putra Malaysia in fulfilment of the requirements for the degree of Doctor of Philosophy January 2015 Abstract of thesis presented to the Senate of Universiti Putra Malaysia in fulfilment of the requirements for the degree of Doctor of Philosophy GENETIC DIVERSITY OF TUBA PLANTS, AND TOXICITY OF THEIR ROTENOIDS FORMULATED AS NANO-EMULSION AGAINST Plutella xylostella L. By NORHAYU BINTI ASIB UPM January 2015 Chairman : Professor Dzolkhifli Omar, PhD. Faculty : Agriculture Rotenone found in the crude extract from the roots of Tuba plants is commonly used as a bioinsecticide to control insect pests of horticultural crops. The control obtained on insect pests varies greatly and this could probably be due to the source and process of extraction, toxicant preparation and application in the field. The need to improve the quality of the rotenone as botanical insecticide and to obtain consistent control of insect pest led to the studies with following objectives; 1) to identify the diversity of Tubaplants based on Random Amplified Polymorphic DNA (RAPD) analysis, Internal Transcribe Spacer (ITS) marker and their morphological characteristics, 2) to extract, and characterize bioactive compound rotenone from Tuba plants, 3) to prepare the bio- based emulsion formulation of rotenone extract and 4) to evaluate the insecticidal properties of the emulsion formulation of rotenone against Diamondback moth (DBM). -

Endosamara Racemosa (Roxb.) Geesink and Callerya Vasta (Kosterm.) Schot
Taiwania, 48(2): 118-128, 2003 Two New Members of the Callerya Group (Fabaceae) Based on Phylogenetic Analysis of rbcL Sequences: Endosamara racemosa (Roxb.) Geesink and Callerya vasta (Kosterm.) Schot (1,3) (1,2) Jer-Ming Hu and Shih-Pai Chang (Manuscript received 2 May, 2003; accepted 29 May, 2003) ABSTRACT: Two new members of Callerya group in Fabaceae, Endosamara racemosa (Roxb.) Geesink and Callerya vasta (Kosterm.) Schot, are identified based on phylogenetic analyses of chloroplast rbcL sequences. These taxa joined with other previously identified taxa in the Callerya group: Afgekia, Callerya, and Wisteria. These genera are resolved as a basal subclade in the Inverted Repeat Lacking Clade (IRLC), which is a large legume group that includes many temperate and herbaceous legumes in the subfamily Papilionoideae, such as Astragalus, Medicago and Pisum, and is not close to other Millettieae. Endosamara is sister to Millettia japonica (Siebold & Zucc.) A. Gray, but only weakly linked with Wisteria and Afgekia. KEY WORDS: Endosamara, Callerya, Millettieae, Millettia, rbcL, Phylogenetic analysis. INTRODUCTION Recent molecular phylogenetic studies of the tribe Millettieae have revealed that the tribe is polyphyletic and several taxa are needed to be segregated from the core Millettieae group. One of the major segregates from Millettieae is the Callerya group, comprising species from Callerya, Wisteria, Afgekia, and Millettia japonica (Siebold & Zucc.) A. Gray. The group is considered to be part of the Inverted-Repeat-Lacking Clade (IRLC; Wojciechowski et al., 1999) including many temperate herbaceous legumes. Such result is consistent and supported by chloroplast inverted repeat surveys (Lavin et al., 1990; Liston, 1995) and phylogenetic studies of the phytochrome gene family (Lavin et al., 1998), chloroplast rbcL (Doyle et al., 1997; Kajita et al., 2001), trnK/matK (Hu et al., 2000), and nuclear ribosomal ITS regions (Hu et al., 2002). -

Ana Cristina Oltramari Toledo.Pdf
UNIVERSIDADE ESTADUAL DE PONTA GROSSA SETOR DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS FARMACÊUTICAS ANA CRISTINA OLTRAMARI TOLEDO DESENVOLVIMENTO, CARACTERIZAÇÃO E AVALIAÇÃO DAS ATIVIDADES BIOLÓGICAS DE NANOPARTÍCULAS DE PRATA E DE OURO, OBTIDAS POR SÍNTESE VERDE, A PARTIR DO EXTRATO AQUOSO DAS SEMENTES DE Pterodon emarginatus Vogel (Fabaceae) ASSOCIADAS À GENTAMICINA E AO ÁCIDO HIALURÔNICO PONTA GROSSA 2021 ANA CRISTINA OLTRAMARI TOLEDO DESENVOLVIMENTO, CARACTERIZAÇÃO E AVALIAÇÃO DAS ATIVIDADES BIOLÓGICAS DE NANOPARTÍCULAS DE PRATA E DE OURO, OBTIDAS POR SÍNTESE VERDE, A PARTIR DO EXTRATO AQUOSO DAS SEMENTES DE Pterodon emarginatus Vogel (Fabaceae) ASSOCIADAS À GENTAMICINA E AO ÁCIDO HIALURÔNICO Tese apresentada para a obtenção do título de doutora na Universidade Estadual de Ponta Grossa, Área de Fármacos, Medicamentos e Biociências Aplicadas à Farmácia. Orientadora: Profa. Dra. Josiane de Fátima Padilha de Paula PONTA GROSSA 2021 Toledo, Ana Cristina Oltramari T649 Desenvolvimento, caracterização e avaliação das atividades biológicas de nanopartículas de prata e de ouro, obtidas por síntese verde, a partir do extrato aquoso das sementes de Pterodon emarginatus Vogel (Fabaceae) associadas à gentamicina e ao ácido hialurônico / Ana Cristina Oltramari Toledo. Ponta Grossa, 2021. 169 f. Tese (Doutorado em Ciências Farmacêuticas - Área de Concentração: Fármacos, Medicamentos e Biociências Aplicadas à Farmácia), Universidade Estadual de Ponta Grossa. Orientadora: Profa. Dra. Josiane de Fátima Padilha de Paula. 1. Pterodon emarginatus Vogel. 2. Síntese verde. 3. Nanopartículas metálicas. 4. Atividades biológicas. I. Paula, Josiane de Fátima Padilha de. II. Universidade Estadual de Ponta Grossa. Fármacos, Medicamentos e Biociências Aplicadas à Farmácia. III.T. CDD: 615.321 Ficha catalográfica elaborada por Maria Luzia Fernandes Bertholino dos Santos- CRB9/986 Programa de Pós-Graduação em Ciências Farmacêuticas Programa de Pós-Graduação em PttOGft"".DliPÓ5-OAfoOU"ÇI.O (11. -

Table of Contents Below) with Family Name Provided
1 Australian Plants Society Plant Table Profiles – Sutherland Group (updated August 2021) Below is a progressive list of all cultivated plants from members’ gardens and Joseph Banks Native Plants Reserve that have made an appearance on the Plant Table at Sutherland Group meetings. Links to websites are provided for the plants so that further research can be done. Plants are grouped in the categories of: Trees and large shrubs (woody plants generally taller than 4 m) Medium to small shrubs (woody plants from 0.1 to 4 m) Ground covers or ground-dwelling (Grasses, orchids, herbaceous and soft-wooded plants, ferns etc), as well as epiphytes (eg: Platycerium) Vines and scramblers Plants are in alphabetical order by botanic names within plants categories (see table of contents below) with family name provided. Common names are included where there is a known common name for the plant: Table of Contents Trees and Large shrubs........................................................................................................................... 2 Medium to small shrubs ...................................................................................................................... 23 Groundcovers and other ground‐dwelling plants as well as epiphytes. ............................................ 64 Vines and Scramblers ........................................................................................................................... 86 Sutherland Group http://sutherland.austplants.com.au 2 Trees and Large shrubs Acacia decurrens -

TESE Ana Rafaela Da Silva Oliveira.Pdf
UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE BIOCIÊNCIAS DEPARTAMENTO DE GENÉTICA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA ANA RAFAELA DA SILVA OLIVEIRA ANÁLISES DE MACROSSINTENIA ENTRE ESPÉCIES DE VIGNA SAVI E DE PHASEOLUS L. MEDIANTE MAPEAMENTO CITOGENÉTICO Recife 2018 2 ANA RAFAELA DA SILVA OLIVEIRA ANÁLISES DE MACROSSINTENIA ENTRE ESPÉCIES DE VIGNA SAVI E DE PHASEOLUS L. MEDIANTE MAPEAMENTO CITOGENÉTICO Tese apresentada ao Programa de Pós-Graduação em Genética da Universidade Federal de Pernambuco como requisito parcial para obtenção do título de Doutora em Genética. Área de concentração: Genética Orientadora: Ana Christina Brasileiro-Vidal Coorientadoras: Ana Maria Benko-Iseppon Andrea Pedrosa-Harand Recife 2018 Catalogação na fonte: Bibliotecária Claudina Queiroz, CRB4/1752 Oliveira, Ana Rafaela da Silva Análises de macrossintenia entre espécies de Vigna savi e de Phaseolus L. Mediante mapeamento citogenético / Ana Rafaela da Silva Oliveira - 2019. 123 folhas: il., fig., tab. Orientadora: Ana Christina Brasileiro Vidal Coorientadoras: Ana Maria Benko Iseppon Andrea Pedrosa Harand Tese (doutorado) – Universidade Federal de Pernambuco. Centro de Biociências. Programa de Pós-Graduação em Genética. Recife, 2019. Inclui referências e anexo 1. Vigna savi 2. Phaseolus L. 3. Mapa cromossômico I. Vidal, Ana Christina Brasileiro (orient.) II. Iseppon, Ana Maria Benko (coorient.) III. Harand, Andrea Pedrosa (coorient.) IV. Título 583.74 CDD (22.ed.) UFPE/CB-2019-096 ANA RAFAELA DA SILVA OLIVEIRA ANÁLISES DE MACROSSINTENIA ENTRE ESPÉCIES DE VIGNA SAVI E DE PHASEOLUS L. MEDIANTE MAPEAMENTO CITOGENÉTICO Tese apresentada ao Programa de Pós- Graduação em Genética da Universidade Federal de Pernambuco, como requisito parcial para a obtenção do título de Doutora em genética. Aprovada em 27/02/2018 BANCA EXAMINADORA: ____________________________________________ Profa. -

Of ODOARDOBECCARI Dedication
1864 1906 1918 Dedicated to the memory of ODOARDOBECCARI Dedication A dedication to ODOARDO BECCARI, the greatest botanist ever to study in Malesia, is long overdue. Although best known as a plant taxonomist, his versatile genius extended far beyond the basic field ofthis branch ofBotany, his wide interest leading him to investigate the laws ofevolution, the interrelations between plants and animals, the connection between vegetation and environ- cultivated ment, plant distribution, the and useful plants of Malesia and many other problems of life. plant But, even if he devoted his studies to plants, in the depth of his mind he was primarily a naturalist, and in his long, lonely and dangerousexplorations in Malesia he was attracted to all aspects ofnature and human life, assembling, besides plants, an incredibly large number of collec- tions and an invaluable wealth ofdrawings and observations in zoology, anthropologyand ethnol- He ogy. was indeed a naturalist, and one of the greatest of his time; but never in his mind were the knowledge and beauty of Nature disjoined, and, as he was a true and complete naturalist, he the time was at same a poet and an artist. His Nelleforestedi Borneo, Viaggi ericerche di un mturalista(1902), excellently translatedinto English (in a somewhatabbreviated form) by Prof. E. GioLiouandrevised and edited by F.H.H. Guillemardas Wanderingsin the great forests of Borneo (1904), is a treasure in tropical botany; it is in fact an unrivalledintroductionto tropical plant lifeand animals, man included. It is a most readable book touching on all sorts of topics and we advise it to be studied by all young people whose ambition it is to devote their life to tropical research. -

Table 7: Species Changing IUCN Red List Status (2018-2019)
IUCN Red List version 2019-3: Table 7 Last Updated: 10 December 2019 Table 7: Species changing IUCN Red List Status (2018-2019) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2018 (IUCN Red List version 2018-2) and 2019 (IUCN Red List version 2019-3) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered [CR(PE) - Critically Endangered (Possibly Extinct), CR(PEW) - Critically Endangered (Possibly Extinct in the Wild)], EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.); E - Previous listing was an Error. IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2018) List (2019) change version Category -

Mt Gravatt Species List - Location Ver 14.1 Plants Family Order
Mt Gravatt Species List - Location ver 14.1 Plants Family Order Weed Family Form Genus Species Common NameFlowering Times Size (approx) Dependent Species Dependent Species Planted Spring Summer Autumn Winter Height Width Amaranthaceae Creeper Weed! Alternanthera nodiflora Common Joyweed Asparagaceae Creeper Weed! Asparagus plumosus Climbing Asparagus Fern Commelinaceae Creeper Weed! Callisia repens Creeping Inch Plant x x Wandering Jew (native), Scurvy Commelinaceae Creeper Commelina diffusa Weed Convolvulaceae Creeper Dichondra repens Kidney weed Devil's Ivy, Pothos, Golden Araceae Creeper Weed! Epipremnum aureum Pothos, Money Plant Fabaceae Creeper Glycine clandestina v clandestina Twining Glycine, Love Creeper Fabaceae Creeper Glycine microphylla Small-leaf Glycine Fabaceae Creeper Glycine tabacina Variable Glycine-pea Fabaceae Creeper Hardenbergia violacea Native Sarsaparilla x Climber Prostrate Common Grass-blue Eastern Spinebill Fabaceae Creeper Kennedia rubicunda Dusky Coral Pea x x Climber Long-tailed Pea-blue Verbenaceae Creeper Weed! Lantana montevidenses Lantana Creeping Fabaceae Creeper Weed! Macroptilium atropurpureum Sirato Fabaceae Creeper Weed! Macrotyloma axillare Perennial Horse Gram Asteraceae Creeper Weed! Sphagneticola trilobata Singapore Daisy Commelinaceae Creeper Weed! Tradescantia albiflora Wandering Jew Commelinaceae Creeper Weed! Tradescantia zebrina Silvery Inch Plant, Zebra Plant Fabaceae Creeper Vigna vexillata var. angustifolia Wild Cow Pea x x Climber Fabaceae Creeper Zornia dyctiocarpa Zornia x 30cm Fabaceae -

Dry Forest Trees of Madagascar
The Red List of Dry Forest Trees of Madagascar Emily Beech, Malin Rivers, Sylvie Andriambololonera, Faranirina Lantoarisoa, Helene Ralimanana, Solofo Rakotoarisoa, Aro Vonjy Ramarosandratana, Megan Barstow, Katharine Davies, Ryan Hills, Kate Marfleet & Vololoniaina Jeannoda Published by Botanic Gardens Conservation International Descanso House, 199 Kew Road, Richmond, Surrey, TW9 3BW, UK. © 2020 Botanic Gardens Conservation International ISBN-10: 978-1-905164-75-2 ISBN-13: 978-1-905164-75-2 Reproduction of any part of the publication for educational, conservation and other non-profit purposes is authorized without prior permission from the copyright holder, provided that the source is fully acknowledged. Reproduction for resale or other commercial purposes is prohibited without prior written permission from the copyright holder. Recommended citation: Beech, E., Rivers, M., Andriambololonera, S., Lantoarisoa, F., Ralimanana, H., Rakotoarisoa, S., Ramarosandratana, A.V., Barstow, M., Davies, K., Hills, BOTANIC GARDENS CONSERVATION INTERNATIONAL (BGCI) R., Marfleet, K. and Jeannoda, V. (2020). Red List of is the world’s largest plant conservation network, comprising more than Dry Forest Trees of Madagascar. BGCI. Richmond, UK. 500 botanic gardens in over 100 countries, and provides the secretariat to AUTHORS the IUCN/SSC Global Tree Specialist Group. BGCI was established in 1987 Sylvie Andriambololonera and and is a registered charity with offices in the UK, US, China and Kenya. Faranirina Lantoarisoa: Missouri Botanical Garden Madagascar Program Helene Ralimanana and Solofo Rakotoarisoa: Kew Madagascar Conservation Centre Aro Vonjy Ramarosandratana: University of Antananarivo (Plant Biology and Ecology Department) THE IUCN/SSC GLOBAL TREE SPECIALIST GROUP (GTSG) forms part of the Species Survival Commission’s network of over 7,000 Emily Beech, Megan Barstow, Katharine Davies, Ryan Hills, Kate Marfleet and Malin Rivers: BGCI volunteers working to stop the loss of plants, animals and their habitats. -
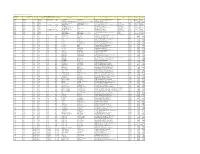
Colgate Palmolive List of Mills As of June 2018 (H1 2018) Direct
Colgate Palmolive List of Mills as of June 2018 (H1 2018) Direct Supplier Second Refiner First Refinery/Aggregator Information Load Port/ Refinery/Aggregator Address Province/ Direct Supplier Supplier Parent Company Refinery/Aggregator Name Mill Company Name Mill Name Country Latitude Longitude Location Location State AgroAmerica Agrocaribe Guatemala Agrocaribe S.A Extractora La Francia Guatemala Extractora Agroaceite Extractora Agroaceite Finca Pensilvania Aldea Los Encuentros, Coatepeque Quetzaltenango. Coatepeque Guatemala 14°33'19.1"N 92°00'20.3"W AgroAmerica Agrocaribe Guatemala Agrocaribe S.A Extractora del Atlantico Guatemala Extractora del Atlantico Extractora del Atlantico km276.5, carretera al Atlantico,Aldea Champona, Morales, izabal Izabal Guatemala 15°35'29.70"N 88°32'40.70"O AgroAmerica Agrocaribe Guatemala Agrocaribe S.A Extractora La Francia Guatemala Extractora La Francia Extractora La Francia km. 243, carretera al Atlantico,Aldea Buena Vista, Morales, izabal Izabal Guatemala 15°28'48.42"N 88°48'6.45" O Oleofinos Oleofinos Mexico Pasternak - - ASOCIACION AGROINDUSTRIAL DE PALMICULTORES DE SABA C.V.Asociacion (ASAPALSA) Agroindustrial de Palmicutores de Saba (ASAPALSA) ALDEA DE ORICA, SABA, COLON Colon HONDURAS 15.54505 -86.180154 Oleofinos Oleofinos Mexico Pasternak - - Cooperativa Agroindustrial de Productores de Palma AceiteraCoopeagropal R.L. (Coopeagropal El Robel R.L.) EL ROBLE, LAUREL, CORREDORES, PUNTARENAS, COSTA RICA Puntarenas Costa Rica 8.4358333 -82.94469444 Oleofinos Oleofinos Mexico Pasternak - - CORPORACIÓN -

Fruits and Seeds of Genera in the Subfamily Faboideae (Fabaceae)
Fruits and Seeds of United States Department of Genera in the Subfamily Agriculture Agricultural Faboideae (Fabaceae) Research Service Technical Bulletin Number 1890 Volume I December 2003 United States Department of Agriculture Fruits and Seeds of Agricultural Research Genera in the Subfamily Service Technical Bulletin Faboideae (Fabaceae) Number 1890 Volume I Joseph H. Kirkbride, Jr., Charles R. Gunn, and Anna L. Weitzman Fruits of A, Centrolobium paraense E.L.R. Tulasne. B, Laburnum anagyroides F.K. Medikus. C, Adesmia boronoides J.D. Hooker. D, Hippocrepis comosa, C. Linnaeus. E, Campylotropis macrocarpa (A.A. von Bunge) A. Rehder. F, Mucuna urens (C. Linnaeus) F.K. Medikus. G, Phaseolus polystachios (C. Linnaeus) N.L. Britton, E.E. Stern, & F. Poggenburg. H, Medicago orbicularis (C. Linnaeus) B. Bartalini. I, Riedeliella graciliflora H.A.T. Harms. J, Medicago arabica (C. Linnaeus) W. Hudson. Kirkbride is a research botanist, U.S. Department of Agriculture, Agricultural Research Service, Systematic Botany and Mycology Laboratory, BARC West Room 304, Building 011A, Beltsville, MD, 20705-2350 (email = [email protected]). Gunn is a botanist (retired) from Brevard, NC (email = [email protected]). Weitzman is a botanist with the Smithsonian Institution, Department of Botany, Washington, DC. Abstract Kirkbride, Joseph H., Jr., Charles R. Gunn, and Anna L radicle junction, Crotalarieae, cuticle, Cytiseae, Weitzman. 2003. Fruits and seeds of genera in the subfamily Dalbergieae, Daleeae, dehiscence, DELTA, Desmodieae, Faboideae (Fabaceae). U. S. Department of Agriculture, Dipteryxeae, distribution, embryo, embryonic axis, en- Technical Bulletin No. 1890, 1,212 pp. docarp, endosperm, epicarp, epicotyl, Euchresteae, Fabeae, fracture line, follicle, funiculus, Galegeae, Genisteae, Technical identification of fruits and seeds of the economi- gynophore, halo, Hedysareae, hilar groove, hilar groove cally important legume plant family (Fabaceae or lips, hilum, Hypocalypteae, hypocotyl, indehiscent, Leguminosae) is often required of U.S.