Studies on the Trichomes of Some Oleaceae, Structure and Ontogeny
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Djvu Document
BULL BQT. ~URV.INDIA Vol. 10, NOS.3 & 4 : pp. 397-400, 1968 TAXONOMIC POSITION OF THE GENUS NYCTANTHES B. C. Kmu AND ANIMADE Bosc Znstifulc, Calcutta ABSTRACT The genus Nyctanthes with only one species N. arbor-tristis Linn. having flowers like that of JaJrninm was originally induded in Oleaceae. In view of its strongly quadrangular stem and its apparent resemblance to Tectona and other members of the family Verbenaceae, Airy Shaw (1952) placed the genus in a new subfamily under the family Verbenaceae. Stant (K952) gave some anatomical evidence for the inclusion of Nyctanthes in the Verbenaceae. On the basis of com- parative study of Nyctanthas, along with some members of Oleaceae, Verbenaceae and Loganiaceae on cytology, general anatomy of stem and leaf, wood anatomy, floral anatomy and palynology and also on the preliminary data of the chemical constituents present in the plants, the authors state that Nyctanfhes has not much affinityto the members of the Verbenaceae, although it has some similarity with several oleaceous members, After taking all points into consideration this genus is assigned to a new family Nyctanthaceae. INTRODUCTION thes is consistent with its being incluiled in the The Genus Nyctanthes Linn. with only .one Verbenaceae. On the basis of anatomical and species N. arbor-tristis Linn. was originally included palynological studies on Nyctanthes arbor-tristis, in the family Oleaceae mainly on account of the Kundu (1966) was of opinion that Nyctamthes structure of the flower which is somewhat like that belongs neither to the Oleaceae nor to the .Verben- of Jasminum. A second species N. -

Verbena Bonariensis
A Horticulture Information article from the Wisconsin Master Gardener website, posted 16 July 2004 Verbena bonariensis Clusters of small purple fl owers fl oating on long, airy stems tempting passing butterfl ies to stop for a sip is just one reason to consider adding Verbena bonairiensis, also called tall verbena or purpletop verbena, to your garden. This plant is just one of about 250 species in the genus Verbena, of which only about half a dozen are in cultivation. V. bonairiensis is native to Brazil and Ar- gentina (the specifi c name is after Buenos Aires). They were fi rst grown as a garden ornamental in 1726 by Englishmen James & William Sherard, who got the seeds from a dried specimen sent back to England from Buenos Aires. This plant is perennial in zones 7 to11 and is grown as an annual in cooler climates. It is considered a weed in many mild climates, such as California, Texas, Australia and southern Africa, where it has naturalized. Verbena bonariensis is a tall, airy plant. Purpletop verbena is an upright, clump-form- ing plant with wiry, widely branched stems. It reaches a height of 3 to 6 feet and spreads 1 to 3 feet. Unlike many other tall herbaceous plants it is unlikely to tip over. The stiff stems and branches are square and rough, like sand paper. The deep green, lance-shaped serrat- ed leaves form a mounded rosette at the base of the plant, with few on the stems. The fl ow- ers are borne in rounded clusters 2 to 3 inches across. -

(Olea Europaea L.) Through Plastome Sequence Comparison Roberto Mariotti1, Nicolò GM Cultrera1, Concepcion Muñoz Díez2, Luciana Baldoni1*, Andrea Rubini1
Mariotti et al. BMC Plant Biology 2010, 10:211 http://www.biomedcentral.com/1471-2229/10/211 RESEARCH ARTICLE Open Access Identification of new polymorphic regions and differentiation of cultivated olives (Olea europaea L.) through plastome sequence comparison Roberto Mariotti1, Nicolò GM Cultrera1, Concepcion Muñoz Díez2, Luciana Baldoni1*, Andrea Rubini1 Abstract Background: The cultivated olive (Olea europaea L.) is the most agriculturally important species of the Oleaceae family. Although many studies have been performed on plastid polymorphisms to evaluate taxonomy, phylogeny and phylogeography of Olea subspecies, only few polymorphic regions discriminating among the agronomically and economically important olive cultivars have been identified. The objective of this study was to sequence the entire plastome of olive and analyze many potential polymorphic regions to develop new inter-cultivar genetic markers. Results: The complete plastid genome of the olive cultivar Frantoio was determined by direct sequence analysis using universal and novel PCR primers designed to amplify all overlapping regions. The chloroplast genome of the olive has an organisation and gene order that is conserved among numerous Angiosperm species and do not contain any of the inversions, gene duplications, insertions, inverted repeat expansions and gene/intron losses that have been found in the chloroplast genomes of the genera Jasminum and Menodora, from the same family as Olea. The annotated sequence was used to evaluate the content of coding genes, the extent, and distribution of repeated and long dispersed sequences and the nucleotide composition pattern. These analyses provided essential information for structural, functional and comparative genomic studies in olive plastids. Furthermore, the alignment of the olive plastome sequence to those of other varieties and species identified 30 new organellar polymorphisms within the cultivated olive. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Medicinal Practices of Sacred Natural Sites: a Socio-Religious Approach for Successful Implementation of Primary
Medicinal practices of sacred natural sites: a socio-religious approach for successful implementation of primary healthcare services Rajasri Ray and Avik Ray Review Correspondence Abstract Rajasri Ray*, Avik Ray Centre for studies in Ethnobiology, Biodiversity and Background: Sacred groves are model systems that Sustainability (CEiBa), Malda - 732103, West have the potential to contribute to rural healthcare Bengal, India owing to their medicinal floral diversity and strong social acceptance. *Corresponding Author: Rajasri Ray; [email protected] Methods: We examined this idea employing ethnomedicinal plants and their application Ethnobotany Research & Applications documented from sacred groves across India. A total 20:34 (2020) of 65 published documents were shortlisted for the Key words: AYUSH; Ethnomedicine; Medicinal plant; preparation of database and statistical analysis. Sacred grove; Spatial fidelity; Tropical diseases Standard ethnobotanical indices and mapping were used to capture the current trend. Background Results: A total of 1247 species from 152 families Human-nature interaction has been long entwined in has been documented for use against eighteen the history of humanity. Apart from deriving natural categories of diseases common in tropical and sub- resources, humans have a deep rooted tradition of tropical landscapes. Though the reported species venerating nature which is extensively observed are clustered around a few widely distributed across continents (Verschuuren 2010). The tradition families, 71% of them are uniquely represented from has attracted attention of researchers and policy- any single biogeographic region. The use of multiple makers for its impact on local ecological and socio- species in treating an ailment, high use value of the economic dynamics. Ethnomedicine that emanated popular plants, and cross-community similarity in from this tradition, deals health issues with nature- disease treatment reflects rich community wisdom to derived resources. -

Antioxidant and Anti-Inflammatory Activities of Nyctanthes Arbor-Tristis Extracts
JOURNAL OF ADVANCED BOTANY AND ZOOLOGY Journal homepage: http://scienceq.org/Journals/JABZ.php Research Article Open Access Antioxidant and Anti-inflammatory Activities Of Nyctanthes Arbor-Tristis Extracts D. Isaac Dhinakaran1*, and G. Sakthivel2 1,2 Department of Biotechnology, Ayya Nadar Janaki Ammal College, Sivakasi 626124,TN, India. *Corresponding author: D. Isaac Dhinakaran, E-mail: [email protected] Received: December 24, 2015, Accepted: February 11, 2016, Published: February 11, 2016. ABSTRACT Oxidative stress plays an important role in chronic complication of diabetes and is postulated to be associated with increased lipid peroxidation. Novel type of antioxidants was identified from Nycanthes arbor-tristi. The antioxidant activity was analyzed for the ethanol extracts of Nyctanthes arbor-Tristis [leaf,flower and stem] by using the DPPH method, total phenol content, Hydrogen peroxide assay, Reducing power assay, and lipid per oxidation assays. It is observed that Nyctanthes arbor-Tristis act as the source of natural antioxidants. Significant anti-inflammatory activity was observed on Nyctanthes arbor-Tristis [leaf,stem,and flower] extracts by using HRBC method. Key words: Nyctanthes arbor-Tristis antioxidant activity, anti-inflammatory activity. INTRODUCTION The methanol extracts of dry flower Nyctanthes Natural product is a source for bioactive compounds and has exhibited high antioxidant activities with high total phenol potential for developing some novel therapeutic agent. Over the content. Phenol compounds such as flavonoids, phenolic acids last decade there has been a growing interest in drugs of plant possess diverse biological properties such as origin and such drugs formed an important class for disease anti-inflammatory, anticarcinogenic and anti-atherosclerotic control.The herbal products today symbolize safety in contrast activities [7] to the synthetics that are regarded as unsafe to human and Nyctanthes leaf extract ameliorates silica induced early environment [1]. -

Olea Europaea L. a Botanical Contribution to Culture
American-Eurasian J. Agric. & Environ. Sci., 2 (4): 382-387, 2007 ISSN 1818-6769 © IDOSI Publications, 2007 Olea europaea L. A Botanical Contribution to Culture Sophia Rhizopoulou National and Kapodistrian University of Athens, Department of Biology, Section of Botany, Panepistimioupoli, Athens 157 84, Greece Abstract: One of the oldest known cultivated plant species is Olea europaea L., the olive tree. The wild olive tree is an evergreen, long-lived species, wide-spread as a native plant in the Mediterranean province. This sacred tree of the goddess Athena is intimately linked with the civilizations which developed around the shores of the Mediterranean and makes a starting point for mythological and symbolic forms, as well as for tradition, cultivation, diet, health and culture. In modern times, the olive has spread widely over the world. Key words: Olea • etymology • origin • cultivation • culture INTRODUCTION Table 1: Classification of Olea ewopaea Superdivi&on Speimatophyta-seed plants Olea europaea L. (Fig. 1 & Table 1) belongs to a Division Magnohophyta-flowenng plants genus of about 20-25 species in the family Oleaceae [1-3] Class Magn olio psi da- dicotyledons and it is one of the earliest cultivated plants. The olive Subclass Astendae- tree is an evergreen, slow-growing species, tolerant to Order Scrophulanale- drought stress and extremely long-lived, with a life Family Oleaceae- olive family expectancy of about 500 years. It is indicative that Genus Olea- olive Species Olea europaea L. -olive Theophrastus, 24 centuries ago, wrote: 'Perhaps we may say that the longest-lived tree is that which in all ways, is able to persist, as does the olive by its trunk, by its power of developing sidegrowth and by the fact its roots are so hard to destroy' [4, book IV. -

Syringa Xchinensis
Syringa xchinensis - Chinese Lilac or Rouen Lilac (Oleaceae) ------------------------------------------------------------------------------------------------------------ Syringa x chinensis is a shrub with showy, early the terminal floral buds, flowering in early- and mid- May, very fragrant inflorescences, but Chinese Lilac May and lasting for 1-2 weeks is susceptible to powdery mildew on its foliage by -inflorescences occur mostly as single-flowering late summer. forms, with relatively few cultivars as compared to Common Lilac FEATURES -while deadheading will slightly improve the overall Form vigor and appearance of the shrub, it is usually -medium-sized to large- impractical to perform except on young shrubs sized ornamental shrub Fruits maturing at about 10' tall x -winter persistent, brown dehiscent capsules occur on 10' wide, but sometimes woody fruiting stalks, not ornamentally effective but larger a good identification feature of the genus -upright oval growth habit Twigs in youth, becoming leggy -light brown to brown-gray, lightly lenticeled, with and spreading with age fairly stout stems having moderately-sized floral buds -medium growth rate (often in pairs at the terminus of the season's growth) Culture and smaller vegetative buds (as lateral buds on the -full sun to partial shade flowering stems, or as lateral and terminal buds on -best performance occurs in full sun in moist, well- portions of the shrub that are more shaded) drained, neutral to slightly acidic soils of average Trunk fertility, in areas with good air circulation; it is highly -multi-trunked, light brown, and slightly exfoliating adaptable to poor soils, soils of various pH, drought, in thin strips with maturity, becoming somewhat and pollution, but declines under the heat and high leggy with age, and with basal suckers having a rapid humidity of the Southern U.S. -

Verbena Bonariensis Tall Vervain
TREATMENT OPTIONS from the book Weed Control in Natural Areas in the Western United States This does not constitute a formal recommendation. When using herbicides always read the label, and when in doubt consult your farm advisor or county agent. This is an excerpt from the book Weed Control in Natural Areas in the Western United States and is available wholesale through the UC Weed Research & Information Center (wric.ucdavis.edu) or retail through the Western Society of Weed Science (wsweedscience.org) or the California Invasive Species Council (cal-ipc.org). Verbena bonariensis Tall vervain Family: Verbenaceae (vervain) NON-CHEMICAL CONTROL Cultural: grazing NIA Cultural: prescribed burning NIA Mechanical: mowing and cutting NIA Mechanical: tillage NIA Mechanical: grubbing, digging or hand pulling NIA CHEMICAL CONTROL The following specific use information is based on published papers and reports by researchers and land managers. Other trade names may be available, and other compounds also are labeled for this weed. Directions for use may vary between brands; see label before use. 2,4-D E Imazapic F* Postemergence Aminocyclopyrachlor + chlorsulfuron NIA Imazapyr E* Aminopyralid NIA Metsulfuron NIA Chlorsulfuron NIA Paraquat NIA Clopyralid NIA Picloram NIA Dicamba NIA Rimsulfuron NIA Glyphosate P─F* Sulfometuron P* Hexazinone P* Sulfosulfuron NIA Triclopyr NIA E = Excellent control, generally better than 95% * = Likely based on results of observations of G = Good control, 80-95% related species FLW = flowering F = Fair control, 50-80% NIA = No information available P = Poor control, below 50% Fa = Fall Control includes effects within the season of treatment. Sp = Spring Control is followed by best timing, if known, when efficacy is E or G. -
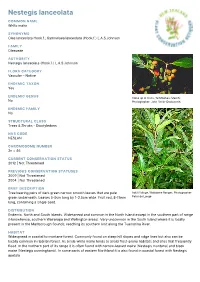
Nestegis Lanceolata
Nestegis lanceolata COMMON NAME White maire SYNONYMS Olea lanceolata Hook.f.; Gymnelaea lanceolata (Hook.f.) L.A.S.Johnson FAMILY Oleaceae AUTHORITY Nestegis lanceolata (Hook.f.) L.A.S.Johnson FLORA CATEGORY Vascular – Native ENDEMIC TAXON Yes ENDEMIC GENUS Close up of fruits, Te Moehau (March). No Photographer: John Smith-Dodsworth ENDEMIC FAMILY No STRUCTURAL CLASS Trees & Shrubs - Dicotyledons NVS CODE NESLAN CHROMOSOME NUMBER 2n = 46 CURRENT CONSERVATION STATUS 2012 | Not Threatened PREVIOUS CONSERVATION STATUSES 2009 | Not Threatened 2004 | Not Threatened BRIEF DESCRIPTION Tree bearing pairs of dark green narrow smooth leaves that are pale Adult foliage, Waitakere Ranges. Photographer: green underneath. Leaves 5-9cm long by 1-2.5cm wide. Fruit red, 8-11mm Peter de Lange long, containing a single seed. DISTRIBUTION Endemic. North and South Islands. Widespread and common in the North Island except in the southern part of range (Horowhenua, southern Wairarapa and Wellington areas). Very uncommon in the South Island where it is locally present in the Marlborough Sounds, reaching its southern limit along the Tuamarina River. HABITAT Widespread in coastal to montane forest. Commonly found on steep hill slopes and ridge lines but also can be locally common in riparian forest. As a rule white maire tends to avoid frost-prone habitats and sites that frequently flood. In the northern part of its range it is often found with narrow-leaved maire (Nestegis montana) and black maire (Nestegis cunninghamii). In some parts of eastern Northland it is also found in coastal forest with Nestegis apetala. FEATURES Stout gynodioecious spreading tree up to 20 m tall usually forming a domed canopy; trunk up to c. -

(Verbenaceae) Using Multiple Loci
bs_bs_banner Botanical Journal of the Linnean Society, 2013, 171, 103–119. With 5 figures Investigating the evolution of Lantaneae (Verbenaceae) using multiple loci PATRICIA LU-IRVING* and RICHARD G. OLMSTEAD Department of Biology, University of Washington, Seattle, Washington 98195, USA Received 3 February 2012; revised 29 June 2012; accepted for publication 23 August 2012 Lantaneae are an example of a taxonomically problematic, widespread and recently radiated Neotropical lineage. Taxonomy in Lantaneae is difficult because of complex, overlapping patterns of shifts in morphological traits among members; monophyly of the traditional genera cannot be assumed without additional information from molecular data. We took a multi-locus approach to infer phylogenetic relationships in Lantaneae, resolving major clades among a broad representative sample that covers the morphological, taxonomic and geographical diversity of this group. Data from multiple, independent loci reveal individual gene trees that are incongruent with one another, with varying degrees of support. Without reliable, applicable methods to determine the sources of such incongru- ence and to resolve it, we present the consensus between well-supported topologies among our data sets as the best estimate of Lantaneae phylogeny to date. According to this consensus tree, fleshy fruits in Lantaneae have been derived from dry fruits at least five times; taxonomic schemes separating genera based on fruit characteristics are artificial. Lantaneae have shifted into the Neotropics from the southern temperate subtropics and have colonized Africa in at least two separate long-distance dispersal events. This study provides a first pass at a broad Lantaneae phylogeny, but two important areas remain unresolved: the position of Acantholippia relative to Aloysia; and species-level relationships in the Lantana–Lippia clade. -

TRADITIONAL MEDICINAL USES, PHYTOCHEMICAL PROFILE and PHARMACOLOGICAL ACTIVITIES of NYCTANTHES ARBORTRIS Hetal Bhalakiya, Nainesh R
Bhalakiya & Modi RJLBPCS 2019 www.rjlbpcs.com Life Science Informatics Publications Original Review Article DOI: 10.26479/2019.0502.76 TRADITIONAL MEDICINAL USES, PHYTOCHEMICAL PROFILE AND PHARMACOLOGICAL ACTIVITIES OF NYCTANTHES ARBORTRIS Hetal Bhalakiya, Nainesh R. Modi* Department of Botany, Bioinformatics and Climate Change Impacts Management, University School of Sciences, Gujarat University, Navrangpura, Ahmedabad, Gujarat, India. ABSTRACT: Nyctanthes arbortristis (Oleaceae) is a mythological plant; and they have high medicinal values in Ayurveda. One of the oldest systems is ayurveda that uses plants and their extracts for treatment and management of various diseases. In India it is considered as one of the most useful conventional medicinal plant. It is considered as an important plant that yields not only unique medicinal products but also has industrial importance. It has several medicinal properties such as anti-helminthic and antipyretic, anti-inflammatory and anti-oxidant activities, hepatoprotective, anti-leishmaniasis, anti-viral, antifungal, anti-pyretic, anti-histaminic, anti- malarial, anti-bacterialbesides it is used as a laxative, in rheumatism, skin ailments and as a sedative. The present review is focus on the potential phytochemicals and pharmacological activity of plant N. arbortristis. Diversity of plant part like seeds, leaves, flowers, bark and fruits have been investigated for their major pharmacological activity and phytochemicals and revealed the presence of flavanoid, glycoside, oleanic acid, essential oils, tannic acid, carotene, fraudulent, lapel, glucose and benzoic acid known for significant hair tonic. KEYWORDS: Nyctanthes arbortristis, Chemical constituents, Pharmacological actions, Bio- prospection. Corresponding Author: Hetal Bhalakiya* Ph.D. Department of Botany, Bioinformatics and Climate Change Impacts Management, University School of Sciences, Gujarat University, Navrangpura, Ahmedabad, Gujarat, India.