Egg Attendance Behaviour of an Amazonian Poison Frog Ameerega Hahneli (Anura: Dendrobatidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Catalogue of the Amphibians of Venezuela: Illustrated and Annotated Species List, Distribution, and Conservation 1,2César L
Mannophryne vulcano, Male carrying tadpoles. El Ávila (Parque Nacional Guairarepano), Distrito Federal. Photo: Jose Vieira. We want to dedicate this work to some outstanding individuals who encouraged us, directly or indirectly, and are no longer with us. They were colleagues and close friends, and their friendship will remain for years to come. César Molina Rodríguez (1960–2015) Erik Arrieta Márquez (1978–2008) Jose Ayarzagüena Sanz (1952–2011) Saúl Gutiérrez Eljuri (1960–2012) Juan Rivero (1923–2014) Luis Scott (1948–2011) Marco Natera Mumaw (1972–2010) Official journal website: Amphibian & Reptile Conservation amphibian-reptile-conservation.org 13(1) [Special Section]: 1–198 (e180). Catalogue of the amphibians of Venezuela: Illustrated and annotated species list, distribution, and conservation 1,2César L. Barrio-Amorós, 3,4Fernando J. M. Rojas-Runjaic, and 5J. Celsa Señaris 1Fundación AndígenA, Apartado Postal 210, Mérida, VENEZUELA 2Current address: Doc Frog Expeditions, Uvita de Osa, COSTA RICA 3Fundación La Salle de Ciencias Naturales, Museo de Historia Natural La Salle, Apartado Postal 1930, Caracas 1010-A, VENEZUELA 4Current address: Pontifícia Universidade Católica do Río Grande do Sul (PUCRS), Laboratório de Sistemática de Vertebrados, Av. Ipiranga 6681, Porto Alegre, RS 90619–900, BRAZIL 5Instituto Venezolano de Investigaciones Científicas, Altos de Pipe, apartado 20632, Caracas 1020, VENEZUELA Abstract.—Presented is an annotated checklist of the amphibians of Venezuela, current as of December 2018. The last comprehensive list (Barrio-Amorós 2009c) included a total of 333 species, while the current catalogue lists 387 species (370 anurans, 10 caecilians, and seven salamanders), including 28 species not yet described or properly identified. Fifty species and four genera are added to the previous list, 25 species are deleted, and 47 experienced nomenclatural changes. -

AC26 Doc. 20 Annex 1 English Only / Únicamente En Inglés / Seulement En Anglais
AC26 Doc. 20 Annex 1 English only / únicamente en inglés / seulement en anglais Fauna: new species and other taxonomic changes relating to species listed in the EC wildlife trade regulations January, 2012 A report to the European Commission Directorate General E - Environment ENV.E.2. – Environmental Agreements and Trade by the United Nations Environment Programme World Conservation Monitoring Centre AC26 Doc. 20, Annex 1 UNEP World Conservation Monitoring Centre 219 Huntingdon Road Cambridge CB3 0DL United Kingdom Tel: +44 (0) 1223 277314 Fax: +44 (0) 1223 277136 Email: [email protected] Website: www.unep-wcmc.org CITATION ABOUT UNEP-WORLD CONSERVATION UNEP-WCMC. 2012. Fauna: new species and MONITORING CENTRE other taxonomic changes relating to species The UNEP World Conservation Monitoring listed in the EC wildlife trade regulations. A Centre (UNEP-WCMC), based in Cambridge, report to the European Commission. UNEP- UK, is the specialist biodiversity information WCMC, Cambridge. and assessment centre of the United Nations Environment Programme (UNEP), run PREPARED FOR cooperatively with WCMC, a UK charity. The Centre's mission is to evaluate and highlight The European Commission, Brussels, Belgium the many values of biodiversity and put authoritative biodiversity knowledge at the DISCLAIMER centre of decision-making. Through the analysis and synthesis of global biodiversity The contents of this report do not necessarily knowledge the Centre provides authoritative, reflect the views or policies of UNEP or strategic and timely information for contributory organisations. The designations conventions, countries and organisations to use employed and the presentations do not imply in the development and implementation of the expressions of any opinion whatsoever on their policies and decisions. -

Species Diversity and Conservation Status of Amphibians in Madre De Dios, Southern Peru
Herpetological Conservation and Biology 4(1):14-29 Submitted: 18 December 2007; Accepted: 4 August 2008 SPECIES DIVERSITY AND CONSERVATION STATUS OF AMPHIBIANS IN MADRE DE DIOS, SOUTHERN PERU 1,2 3 4,5 RUDOLF VON MAY , KAREN SIU-TING , JENNIFER M. JACOBS , MARGARITA MEDINA- 3 6 3,7 1 MÜLLER , GIUSEPPE GAGLIARDI , LILY O. RODRÍGUEZ , AND MAUREEN A. DONNELLY 1 Department of Biological Sciences, Florida International University, 11200 SW 8th Street, OE-167, Miami, Florida 33199, USA 2 Corresponding author, e-mail: [email protected] 3 Departamento de Herpetología, Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Avenida Arenales 1256, Lima 11, Perú 4 Department of Biology, San Francisco State University, 1600 Holloway Avenue, San Francisco, California 94132, USA 5 Department of Entomology, California Academy of Sciences, 55 Music Concourse Drive, San Francisco, California 94118, USA 6 Departamento de Herpetología, Museo de Zoología de la Universidad Nacional de la Amazonía Peruana, Pebas 5ta cuadra, Iquitos, Perú 7 Programa de Desarrollo Rural Sostenible, Cooperación Técnica Alemana – GTZ, Calle Diecisiete 355, Lima 27, Perú ABSTRACT.—This study focuses on amphibian species diversity in the lowland Amazonian rainforest of southern Peru, and on the importance of protected and non-protected areas for maintaining amphibian assemblages in this region. We compared species lists from nine sites in the Madre de Dios region, five of which are in nationally recognized protected areas and four are outside the country’s protected area system. Los Amigos, occurring outside the protected area system, is the most species-rich locality included in our comparison. -

Chromosome Analysis of Five Brazilian
c Indian Academy of Sciences RESEARCH ARTICLE Chromosome analysis of five Brazilian species of poison frogs (Anura: Dendrobatidae) PAULA CAMARGO RODRIGUES1, ODAIR AGUIAR2, FLÁVIA SERPIERI1, ALBERTINA PIMENTEL LIMA3, MASAO UETANEBARO4 and SHIRLEI MARIA RECCO-PIMENTEL1∗ 1Departamento de Anatomia, Biologia Celular e Fisiologia, Instituto de Biologia, Universidade Estadual de Campinas, 13083-863 Campinas, São Paulo, Brazil 2Departamento de Biociências, Universidade Federal de São Paulo, Campus Baixada Santista, 11060-001 Santos, São Paulo, Brazil 3Coordenadoria de Pesquisas em Ecologia, Instituto Nacional de Pesquisas do Amazonas, 69011-970 Manaus, Amazonas, Brazil 4Departamento de Biologia, Universidade Federal de Mato Grosso do Sul, 70070-900 Campo Grande, Mato Grosso do Sul, Brazil Abstract Dendrobatid frogs have undergone an extensive systematic reorganization based on recent molecular findings. The present work describes karyotypes of the Brazilian species Adelphobates castaneoticus, A. quinquevittatus, Ameerega picta, A. galactonotus and Dendrobates tinctorius which were compared to each other and with previously described related species. All karyotypes consisted of 2n = 18 chromosomes, except for A. picta which had 2n = 24. The karyotypes of the Adelphobates and D. tinctorius species were highly similar to each other and to the other 2n = 18 previously studied species, revealing conserved karyotypic characteristics in both genera. In recent phylogenetic studies, all Adelphobates species were grouped in a clade separated from the Dendrobates species. Thus, we hypothesized that their common karyotypic traits may have a distinct origin by chromosome rearrangements and mutations. In A. picta, with 2n = 24, chromosome features of pairs from 1 to 8 are shared with other previously karyotyped species within this genus. Hence, the A. -
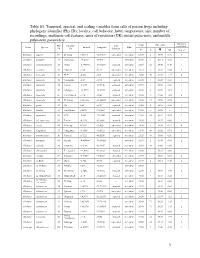
1 Table S1. Temporal, Spectral, and Scaling Variables from Calls Of
Table S1. Temporal, spectral, and scaling variables from calls of poison frogs including phylogeny identifier (Phy ID), locality, call behavior, habit, temperature, size, number of recordings, multinote call features, units of repetition (UR), initial pulse-note, and middle pulse-note parameters. Analyzed Phy Locality Call Temp SVL (mm) Genus Species Latitude Longitude Habit recordings ID ID Behavior °C N ! SD N of ♂ Allobates algorei 60 El Tama 7.65375 -72.19137 concealed terrestrial 23.50 8 18.90 0.70 3 Allobates brunneus 37 Guimaraes -15.2667 -55.5311 -- terrestrial 26.50 1 16.13 0.00 1 Allobates caeruleodactylus 48 Borba -4.398593 -59.60251 exposed terrestrial 25.60 12 15.50 0.40 1 Allobates crombiei 52 Altamira -3.65 -52.38 concealed terrestrial 24.10 2 18.10 0.04 2 Allobates femoralis 43 ECY -0.633 -76.5 concealed terrestrial 25.60 20 23.58 1.27 6 Allobates femoralis 46 Porongaba -8.67 -72.78 exposed terrestrial 25.00 1 25.38 0.00 1 Allobates femoralis 44 Leticia -4.2153 -69.9406 exposed terrestrial 25.50 1 20.90 0.00 1 Allobates femoralis 40 Albergue -12.8773 -71.3865 exposed terrestrial 26.00 6 21.98 2.18 1 Allobates femoralis 41 CAmazonico -12.6 -70.08 exposed terrestrial 26.00 12 22.43 1.06 4 Allobates femoralis 45 El Palmar 8.333333 -61.66667 concealed terrestrial 24.00 27 25.50 0.76 1 Allobates granti 49 FG 3.62 -53.17 exposed terrestrial 24.60 8 16.15 0.55 1 Allobates humilis 59 San Ramon 8.8678 -70.4861 concealed terrestrial 19.50 -- 21.80 -- 1 Allobates insperatus 54 ECY -0.633 -76.4005 exposed terrestrial 24.60 18 16.64 0.93 7 Allobates aff. -

Taxonomic Checklist of Amphibian Species Listed in the CITES
CoP17 Doc. 81.1 Annex 5 (English only / Únicamente en inglés / Seulement en anglais) Taxonomic Checklist of Amphibian Species listed in the CITES Appendices and the Annexes of EC Regulation 338/97 Species information extracted from FROST, D. R. (2015) "Amphibian Species of the World, an online Reference" V. 6.0 (as of May 2015) Copyright © 1998-2015, Darrel Frost and TheAmericanMuseum of Natural History. All Rights Reserved. Additional comments included by the Nomenclature Specialist of the CITES Animals Committee (indicated by "NC comment") Reproduction for commercial purposes prohibited. CoP17 Doc. 81.1 Annex 5 - p. 1 Amphibian Species covered by this Checklist listed by listed by CITES EC- as well as Family Species Regulation EC 338/97 Regulation only 338/97 ANURA Aromobatidae Allobates femoralis X Aromobatidae Allobates hodli X Aromobatidae Allobates myersi X Aromobatidae Allobates zaparo X Aromobatidae Anomaloglossus rufulus X Bufonidae Altiphrynoides malcolmi X Bufonidae Altiphrynoides osgoodi X Bufonidae Amietophrynus channingi X Bufonidae Amietophrynus superciliaris X Bufonidae Atelopus zeteki X Bufonidae Incilius periglenes X Bufonidae Nectophrynoides asperginis X Bufonidae Nectophrynoides cryptus X Bufonidae Nectophrynoides frontierei X Bufonidae Nectophrynoides laevis X Bufonidae Nectophrynoides laticeps X Bufonidae Nectophrynoides minutus X Bufonidae Nectophrynoides paulae X Bufonidae Nectophrynoides poyntoni X Bufonidae Nectophrynoides pseudotornieri X Bufonidae Nectophrynoides tornieri X Bufonidae Nectophrynoides vestergaardi -

Reproductive Biology of Ameerega Trivittata(Anura: Dendrobatidae)
ACTA AMAZONICA http://dx.doi.org/10.1590/1809-4392201305384 Reproductive biology of Ameerega trivittata (Anura: Dendrobatidae) in an area of terra firme forest in eastern Amazonia Ellen Cristina Serrão ACIOLI1 * , Selvino NECKEL-OLIVEIRA2 ¹ Universidade Federal do Pará/Museu Paraense Emílio Goeldi, Programa de Pós Graduação em Zoologia, Av. Perimetral 1901/1907, Terra Firme, 66017-970, Belém, Pará, Brasil. ² Universidade Federal de Santa Catarina, Departamento de Ecologia e Zoologia, Centro de Ciências Biológicas, 88040-970, Florianópolis, Santa Catarina, Brasil. * Corresponding author: [email protected] ABSTRACT The reproductive success of tropical amphibians is influenced by factors such as body size and the characteristics of breeding sites. Data on reproductive biology are important for the understanding of population dynamics and the maintenance of species. The objectives of the present study were to examine the abundance ofAmeerega trivittata, analyze the use of microhabitats by calling males and the snout-vent length (SVL) of breeding males and females, the number of tadpoles carried by the males and mature oocytes in the females, as well as the relationship between the SVL of the female and both the number and mean size of the mature oocytes found in the ovaries. Three field trips were conducted between January and September, 2009. A total of 31 plots, with a mean area of 2.3 ha, were surveyed, resulting in records of 235 individuals, with a mean density of 3.26 individuals per hectare. Overall, 66.1% of the individuals sighted were located in the leaf litter, while 17.4% were perched on decaying tree trunks on the forest floor, 15.7% on the aerial roots ofCecropia trees, and 0.8% on lianas. -

Diet Composition of Ameerega Picta
Bonn zoological Bulletin 68 (1): 93–96 ISSN 2190–7307 2019 · Landgref Filho P. et al. http://www.zoologicalbulletin.de https://doi.org/10.20363/BZB-2019.68.1.093 Scientific note urn:lsid:zoobank.org:pub:CBA156F4-E9AA-401C-B036-77C362CE1E89 Diet composition of Ameerega picta (Tschudi, 1838) from the Serra da Bodoquena region in central Brazil, with a summary of dietary studies on species of the genus Ameerega (Anura: Dendrobatidae) Paulo Landgref Filho1, Fabrício H. Oda2, *, Fabio T. Mise3, Domingos de J. Rodrigues4 & Masao Uetanabaro5 1 Universidade Federal de Mato Grosso do Sul, Campus Aquidauana, 79200-000, Aquidauana, Mato Grosso do Sul, Brazil 2 Departamento de Química Biológica, Programa de Pós-graduação em Bioprospecção Molecular, Universidade Regional do Cariri, 63105-00, Crato, Ceará, Brazil 3 Departamento de Biologia, Universidade Estadual do Centro-Oeste, 85040-080, Guarapuava, Paraná, Brazil 4 Instituto de Ciências Naturais, Humanas e Sociais, Universidade Federal de Mato Grosso – Campus Universitário de Sinop, 78557-267, Sinop, Mato Grosso, Brazil 4 Instituto Nacional de Ciência e Tecnologia de Estudos Integrados da Biodiversidade Amazônica – Núcleo Regional de Sinop, Sinop, Mato Grosso, Brazil 5 Rua Clóvis n. 24, 79022-071, Campo Grande, Mato Grosso do Sul, Brazil * Corresponding author: Email: [email protected] 1 urn:lsid:zoobank.org:author:5821C96D-4D9E-4E1D-BE9B-C231B77D1AF3 2 urn:lsid:zoobank.org:author:349712B5-E1B2-4059-A826-58F081DD3A4D 3 urn:lsid:zoobank.org:author:E996AF2A-6CB7-4B73-8DEF-AB7C5EEE1AA0 4 urn:lsid:zoobank.org:author:ACFBA432-3220-4910-AFF5-8FF85053A872 5 urn:lsid:zoobank.org:author:626F6AE3-7D90-4500-BFDC-4E6F4E01B378 Abstract. -

Twenty-Fifth Meeting of the Animals Committee
AC25 Doc. 22 (Rev. 1) Annex 3 (English only / únicamente en inglés / seulement en anglais) Annex 3 Fauna: new species and other changes relating to species listed in the EC wildlife trade regulations – Report compiled by UNEP-WCMC to the European Commission, March, 2011 AC25 Doc. 22 (Rev. 1) Annex 3 – p. 1 Fauna: new species and other taxonomic changes relating to species listed in the EC wildlife trade regulations March, 2011 A report to the European Commission Directorate General E - Environment ENV.E.2. – Environmental Agreements and Trade by the United Nations Environment Programme World Conservation Monitoring Centre AC25 Doc. 22 (Rev. 1) Annex 3 – p. 2 UNEP World Conservation Monitoring Centre 219 Huntingdon Road Cambridge CB3 0DL United Kingdom Tel: +44 (0) 1223 277314 Fax: +44 (0) 1223 277136 Email: [email protected] Website: www.unep-wcmc.org CITATION UNEP-WCMC. 2011. Fauna: new species and other taxonomic changes relating to species ABOUT UNEP-WORLD CONSERVATION listed in the EC wildlife trade regulations. A MONITORING CENTRE report to the European Commission. UNEP- The UNEP World Conservation Monitoring WCMC, Cambridge. Centre (UNEP-WCMC), based in Cambridge, UK, is the specialist biodiversity information and assessment centre of the United Nations Environment Programme (UNEP), run PREPARED FOR cooperatively with WCMC, a UK charity. The The European Commission, Brussels, Belgium Centre's mission is to evaluate and highlight the many values of biodiversity and put authoritative biodiversity knowledge at the DISCLAIMER centre of decision-making. Through the analysis and synthesis of global biodiversity knowledge The contents of this report do not necessarily the Centre provides authoritative, strategic and reflect the views or policies of UNEP or timely information for conventions, countries contributory organisations. -

ANFIBIOS De Cordillera Azul
Parque Nacional Cordillera Azul, San Martín, Loreto, Ucayali y Huánuco, PERÚ 1 ANFIBIOS de Cordillera Azul Kaitlin Tasker y Evan Twomey Fotógrafo indicado debajo de cada foto. Producido por: Tyana Wachter, R. Foster, J. Philipp. Con el apoyo de Connie Keller, el Keller Action Science Center y el Andrew W. Mellon Foundation. © Kaitlin Tasker [[email protected]] y Evan Twomey [[email protected]] y William Duellman [[email protected]] (M): Macho, (H): Hembra, (Juv.): Juvenil © Science and Education, The Field Museum, Chicago [[email protected]] [fieldguides.fieldmuseum.org] Rapid Color Guide # 596 versión 1 10/2015 1 Allobates femoralis 2 Allobates femoralis 3 Allobates ornatus 4 Allobates trilineatus AROMOBATIDAE AROMOBATIDAE AROMOBATIDAE AROMOBATIDAE (foto: K. Tasker) (foto: W. Duellman) (foto: W. Duellman) (foto: W. Duellman) 5 Atelopus pulcher 6 Atelopus pulcher 7 Rhinella cf. festae 8 Rhinella cf. festae BUFONIDAE BUFONIDAE BUFONIDAE BUFONIDAE (foto: K. Tasker) (foto: K. Tasker) (foto: K. Tasker) (foto: K. Tasker) 9 Rhinella cf. festae 10 Rhinella margaritifera 11 Rhinella margaritifera 12 Rhinella marina BUFONIDAE BUFONIDAE BUFONIDAE BUFONIDAE (foto: K. Tasker) (foto: K. Tasker) (foto: K. Tasker) 13 Rhinella marina 14 Rhinella poeppigii 15 Cochranella erminea 16 Hyalinobatrachium cf. carlesvilai BUFONIDAE BUFONIDAE CENTROLENIDAE CENTROLENIDAE (foto: K. Tasker) (foto: W. Duellman) (foto: E. Twomey) (foto: E. Twomey) 17 Hyalinobatrachium pellucidum 18 Hyalinobatrachium pellucidum 19 Hyalinobatrachium pellucidum 20 Nymphargus chancas CENTROLENIDAE CENTROLENIDAE CENTROLENIDAE CENTROLENIDAE (foto: W. Duellman) (foto: W. Duellman) (foto: K. Tasker) (foto: E. Twomey) Parque Nacional Cordillera Azul, San Martín, Loreto, Ucayali y Huánuco, PERÚ 2 ANFIBIOS de Cordillera Azul Kaitlin Tasker y Evan Twomey Fotógrafo indicado debajo de cada foto. -

Does Batrachotoxin Autoresistance Co-Evolve with Toxicity in Phyllobates Poison-Dart Frogs?
Does batrachotoxin autoresistance co-evolve with toxicity in Phyllobates poison-dart frogs? Supplementary material Supplementary Tables Table S1. Specimen information and accession numbers of the sequences used in ancestral sequence reconstructions. For sequences obtained from publicly available genomes or transcriptomes where sequences do not have individual accession numbers (i.e. Nanorana parkeri, Rhinella marina, Rana pipiens) we provide the name of the contig or gene annotation. Species Voucher Locality DI DII DIII DIV Source Observations Allobates femoralis- Lumbaquí, Ecuador KT989177 - - KT989148 Tarvin et al. 2016 Allobates talamancae - Tundaloma, Ecuador KT989178 - - KT989149 Tarvin et al. 2016 Allobates zaparo- Río Pastaza, Ecuador KT989179 - - KT989150 Tarvin et al. 2016 Ameerega bilinguis- Venecia, Ecuador KT989180 KT989205 KT989220 KT989151 Tarvin et al. 2016 Ameerega hahneli- Canelos, Ecuador KT989181 - - KT989152 Tarvin et al. 2016 Ameerega parvula- Gualaquiza, Ecuador KT989182 KT989206 - KT989153 Tarvin et al. 2016 Panguana Biological Ameerega petersi GECOH1407 Station, Puerto Inca, Perú TBA - - TBA This Study Panguana Biological Ameerega picta GECOH1406 Station, Puerto Inca, Perú TBA - - TBA This Study Leticia, Amazonas, Ameerega trivittata TNHC-FS466 Colombia TBA - - TBA This Study La Celia, Risaralda, Andinobates bombetes GECOH1495 Colombia TBA - - TBA This Study Yotoco, Valle del Cauca, Andinobates bombetes GECOH297 Colombia TBA - - TBA This Study Andinobates fulguritus GECOH1314 Lloró, Chocó, Colombia TBA - - TBA This -

The Amphibians of the Mitaraka Massif, French Guiana
DIRECTEUR DE LA PUBLICATION : Bruno David Président du Muséum national d’Histoire naturelle RÉDACTRICE EN CHEF / EDITOR-IN-CHIEF : Laure Desutter-Grandcolas ASSISTANTS DE RÉDACTION / ASSISTANT EDITORS : Anne Mabille ([email protected]), Emmanuel Côtez MISE EN PAGE / PAGE LAYOUT : Anne Mabille COMITÉ SCIENTIFIQUE / SCIENTIFIC BOARD : James Carpenter (AMNH, New York, États-Unis) Maria Marta Cigliano (Museo de La Plata, La Plata, Argentine) Henrik Enghoff (NHMD, Copenhague, Danemark) Rafael Marquez (CSIC, Madrid, Espagne) Peter Ng (University of Singapore) Norman I. Platnick (AMNH, New York, États-Unis) Jean-Yves Rasplus (INRA, Montferrier-sur-Lez, France) Jean-François Silvain (IRD, Gif-sur-Yvette, France) Wanda M. Weiner (Polish Academy of Sciences, Cracovie, Pologne) John Wenzel (The Ohio State University, Columbus, États-Unis) COUVERTURE / COVER : Flooded bank of Alama river in the Mitaraka Mountains (French Guiana) (photo Marc Pollet). In medaillon, Synapturanus cf. mirandaribeiroi Nelson & Lescure, 1975. Zoosystema est indexé dans / Zoosystema is indexed in: – Science Citation Index Expanded (SciSearch®) – ISI Alerting Services® – Current Contents® / Agriculture, Biology, and Environmental Sciences® – Scopus® Zoosystema est distribué en version électronique par / Zoosystema is distributed electronically by: – BioOne® (http://www.bioone.org) Les articles ainsi que les nouveautés nomenclaturales publiés dans Zoosystema sont référencés par / Articles and nomenclatural novelties published in Zoosystema are referenced by: – ZooBank® (http://zoobank.org) Zoosystema est une revue en flux continu publiée par les Publications scientifiques du Muséum, Paris / Zoosystema is a fast track journal published by the Museum Science Press, Paris Les Publications scientifiques du Muséum publient aussi / The Museum Science Press also publish: Adansonia, Geodiversitas, Anthropozoologica, European Journal of Taxonomy, Naturae, Cryptogamie sous-sections Algologie, Bryologie, Mycologie.