Predation on the Three-Striped Poison Frog, Ameerega Trivitatta
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Species Diversity and Conservation Status of Amphibians in Madre De Dios, Southern Peru
Herpetological Conservation and Biology 4(1):14-29 Submitted: 18 December 2007; Accepted: 4 August 2008 SPECIES DIVERSITY AND CONSERVATION STATUS OF AMPHIBIANS IN MADRE DE DIOS, SOUTHERN PERU 1,2 3 4,5 RUDOLF VON MAY , KAREN SIU-TING , JENNIFER M. JACOBS , MARGARITA MEDINA- 3 6 3,7 1 MÜLLER , GIUSEPPE GAGLIARDI , LILY O. RODRÍGUEZ , AND MAUREEN A. DONNELLY 1 Department of Biological Sciences, Florida International University, 11200 SW 8th Street, OE-167, Miami, Florida 33199, USA 2 Corresponding author, e-mail: [email protected] 3 Departamento de Herpetología, Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Avenida Arenales 1256, Lima 11, Perú 4 Department of Biology, San Francisco State University, 1600 Holloway Avenue, San Francisco, California 94132, USA 5 Department of Entomology, California Academy of Sciences, 55 Music Concourse Drive, San Francisco, California 94118, USA 6 Departamento de Herpetología, Museo de Zoología de la Universidad Nacional de la Amazonía Peruana, Pebas 5ta cuadra, Iquitos, Perú 7 Programa de Desarrollo Rural Sostenible, Cooperación Técnica Alemana – GTZ, Calle Diecisiete 355, Lima 27, Perú ABSTRACT.—This study focuses on amphibian species diversity in the lowland Amazonian rainforest of southern Peru, and on the importance of protected and non-protected areas for maintaining amphibian assemblages in this region. We compared species lists from nine sites in the Madre de Dios region, five of which are in nationally recognized protected areas and four are outside the country’s protected area system. Los Amigos, occurring outside the protected area system, is the most species-rich locality included in our comparison. -

Thermal Adaptation of Amphibians in Tropical Mountains
Thermal adaptation of amphibians in tropical mountains. Consequences of global warming Adaptaciones térmicas de anfibios en montañas tropicales: consecuencias del calentamiento global Adaptacions tèrmiques d'amfibis en muntanyes tropicals: conseqüències de l'escalfament global Pol Pintanel Costa ADVERTIMENT. La consulta d’aquesta tesi queda condicionada a l’acceptació de les següents condicions d'ús: La difusió d’aquesta tesi per mitjà del servei TDX (www.tdx.cat) i a través del Dipòsit Digital de la UB (diposit.ub.edu) ha estat autoritzada pels titulars dels drets de propietat intel·lectual únicament per a usos privats emmarcats en activitats d’investigació i docència. No s’autoritza la seva reproducció amb finalitats de lucre ni la seva difusió i posada a disposició des d’un lloc aliè al servei TDX ni al Dipòsit Digital de la UB. No s’autoritza la presentació del seu contingut en una finestra o marc aliè a TDX o al Dipòsit Digital de la UB (framing). Aquesta reserva de drets afecta tant al resum de presentació de la tesi com als seus continguts. En la utilització o cita de parts de la tesi és obligat indicar el nom de la persona autora. ADVERTENCIA. La consulta de esta tesis queda condicionada a la aceptación de las siguientes condiciones de uso: La difusión de esta tesis por medio del servicio TDR (www.tdx.cat) y a través del Repositorio Digital de la UB (diposit.ub.edu) ha sido autorizada por los titulares de los derechos de propiedad intelectual únicamente para usos privados enmarcados en actividades de investigación y docencia. -

N.Orntates PUBLISHED by the AMERICAN MUSEUM of NATURAL HISTORY CENTRAL PARK WEST at 79TH STREET, NEW YORK, N.Y
AMERICAN MUSEUM N.orntates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 3068, 15 pp., 12 figures, 1 table June 11, 1993 A New Poison Frog from Manu INational Park, Southeastern Peru (Dendrobatidae, Epipedobates) LILY RODRIGUEZ' AND CHARLES W. MYERS2 ABSTRACT Epipedobates macero is a new species of den- ilar to a few other species occurring along the An- drobatid poison frog from lowland rain forest of dean front in eastern Peru, namely E. petersi and the Manu National Park, in the upper Madre de E. cainarachi, which differ in details ofcoloration, Dios drainage ofsoutheastern Peru. It is most sim- morphology, and vocalization. RESUMEN Epipedobates macero, especie nueva, es un den- del llano amaz6nico al pie de los Andes orientales drobatido venenoso de la selva pluvial baja del peruanos, a saber, E. petersi y E. cainarachi, las Parque Nacional del Manu, en el drenaje del Rio cuales difieren en detalles de coloracion, morfo- Alto Madre de Dios, al sudeste del Peru. Es similar logla, y vocalizacion. a otras dos especies que ocurren en los bosques ' Field Associate, Department of Herpetology and Ichthyology, American Museum of Natural History. Investi- gadora Asociada: Asociaci6n Peruana para la Conservaci6n de la Naturaleza (APECO), Parque Jos6 de Acosta 187, Lima 17, Perfi; and Museo de Historia Natural de la Universidad Mayor de San Marcos, apartado 140434, Lima 14, Peru. 2 Curator, Department of Herpetology and Ichthyology, American Museum of Natural History. Copyright © American Museum of Natural History 1993 ISSN 0003-0082 / Price $3.90 2 AMERICAN MUSEUM NOVITATES NO. -

Cop18 Doc. 62 (Rev
Original language: Spanish CoP18 Doc. 62 (Rev. 1) CONVENTION ON INTERNATIONAL TRADE IN ENDANGERED SPECIES OF WILD FAUNA AND FLORA ____________________ Eighteenth meeting of the Conference of the Parties Colombo (Sri Lanka), 23 May – 3 June 2019 Species specific matters DRAFT DECISIONS ON THE CONSERVATION OF AMPHIBIANS (AMPHIBIA) 1. This document has been submitted by Costa Rica.* 2. Amphibians are the most threatened class of vertebrates worldwide. According to the IUCN Red List of Threatened Species, "of the 6260 amphibian species assessed, nearly one-third of species (32.4 %) are globally threatened or extinct, representing 2030 species". This is considerably higher than the comparable figure for birds (13 %) or mammals (22 %) (IUCN Red List 2016). The number of species classified as Critically Endangered, Endangered, or Vulnerable has increased throughout the world due to a number of threats, including habitat loss, trade, and disease. While loss of habitat and disease are considered to be the major threats to amphibian populations, harvesting for human use is a further pressure, sometimes posing the greatest threat (IUCN, 2016). 3. Amphibians occur throughout the world, with diversity nuclei in Central America, South America, East and Central Africa, East and South Asia, and Madagascar (Abraham et al. 2013; Jenkins et al. 2013; Pratihar et al. 2014; Pimm et al. 2014, among other authors). Amphibians live in a variety of terrestrial and freshwater ecosystems, ranging from tropical forests to deserts (Stuart et al. 2008). 4. Amphibians have been an important part of human culture for millennia and traditionally served as a source of food (e.g., Mohneke et al. -

Chromosome Analysis of Five Brazilian
c Indian Academy of Sciences RESEARCH ARTICLE Chromosome analysis of five Brazilian species of poison frogs (Anura: Dendrobatidae) PAULA CAMARGO RODRIGUES1, ODAIR AGUIAR2, FLÁVIA SERPIERI1, ALBERTINA PIMENTEL LIMA3, MASAO UETANEBARO4 and SHIRLEI MARIA RECCO-PIMENTEL1∗ 1Departamento de Anatomia, Biologia Celular e Fisiologia, Instituto de Biologia, Universidade Estadual de Campinas, 13083-863 Campinas, São Paulo, Brazil 2Departamento de Biociências, Universidade Federal de São Paulo, Campus Baixada Santista, 11060-001 Santos, São Paulo, Brazil 3Coordenadoria de Pesquisas em Ecologia, Instituto Nacional de Pesquisas do Amazonas, 69011-970 Manaus, Amazonas, Brazil 4Departamento de Biologia, Universidade Federal de Mato Grosso do Sul, 70070-900 Campo Grande, Mato Grosso do Sul, Brazil Abstract Dendrobatid frogs have undergone an extensive systematic reorganization based on recent molecular findings. The present work describes karyotypes of the Brazilian species Adelphobates castaneoticus, A. quinquevittatus, Ameerega picta, A. galactonotus and Dendrobates tinctorius which were compared to each other and with previously described related species. All karyotypes consisted of 2n = 18 chromosomes, except for A. picta which had 2n = 24. The karyotypes of the Adelphobates and D. tinctorius species were highly similar to each other and to the other 2n = 18 previously studied species, revealing conserved karyotypic characteristics in both genera. In recent phylogenetic studies, all Adelphobates species were grouped in a clade separated from the Dendrobates species. Thus, we hypothesized that their common karyotypic traits may have a distinct origin by chromosome rearrangements and mutations. In A. picta, with 2n = 24, chromosome features of pairs from 1 to 8 are shared with other previously karyotyped species within this genus. Hence, the A. -
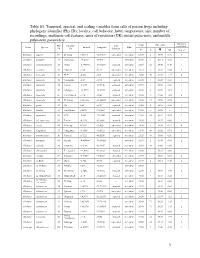
1 Table S1. Temporal, Spectral, and Scaling Variables from Calls Of
Table S1. Temporal, spectral, and scaling variables from calls of poison frogs including phylogeny identifier (Phy ID), locality, call behavior, habit, temperature, size, number of recordings, multinote call features, units of repetition (UR), initial pulse-note, and middle pulse-note parameters. Analyzed Phy Locality Call Temp SVL (mm) Genus Species Latitude Longitude Habit recordings ID ID Behavior °C N ! SD N of ♂ Allobates algorei 60 El Tama 7.65375 -72.19137 concealed terrestrial 23.50 8 18.90 0.70 3 Allobates brunneus 37 Guimaraes -15.2667 -55.5311 -- terrestrial 26.50 1 16.13 0.00 1 Allobates caeruleodactylus 48 Borba -4.398593 -59.60251 exposed terrestrial 25.60 12 15.50 0.40 1 Allobates crombiei 52 Altamira -3.65 -52.38 concealed terrestrial 24.10 2 18.10 0.04 2 Allobates femoralis 43 ECY -0.633 -76.5 concealed terrestrial 25.60 20 23.58 1.27 6 Allobates femoralis 46 Porongaba -8.67 -72.78 exposed terrestrial 25.00 1 25.38 0.00 1 Allobates femoralis 44 Leticia -4.2153 -69.9406 exposed terrestrial 25.50 1 20.90 0.00 1 Allobates femoralis 40 Albergue -12.8773 -71.3865 exposed terrestrial 26.00 6 21.98 2.18 1 Allobates femoralis 41 CAmazonico -12.6 -70.08 exposed terrestrial 26.00 12 22.43 1.06 4 Allobates femoralis 45 El Palmar 8.333333 -61.66667 concealed terrestrial 24.00 27 25.50 0.76 1 Allobates granti 49 FG 3.62 -53.17 exposed terrestrial 24.60 8 16.15 0.55 1 Allobates humilis 59 San Ramon 8.8678 -70.4861 concealed terrestrial 19.50 -- 21.80 -- 1 Allobates insperatus 54 ECY -0.633 -76.4005 exposed terrestrial 24.60 18 16.64 0.93 7 Allobates aff. -

Taxonomic Checklist of Amphibian Species Listed in the CITES
CoP17 Doc. 81.1 Annex 5 (English only / Únicamente en inglés / Seulement en anglais) Taxonomic Checklist of Amphibian Species listed in the CITES Appendices and the Annexes of EC Regulation 338/97 Species information extracted from FROST, D. R. (2015) "Amphibian Species of the World, an online Reference" V. 6.0 (as of May 2015) Copyright © 1998-2015, Darrel Frost and TheAmericanMuseum of Natural History. All Rights Reserved. Additional comments included by the Nomenclature Specialist of the CITES Animals Committee (indicated by "NC comment") Reproduction for commercial purposes prohibited. CoP17 Doc. 81.1 Annex 5 - p. 1 Amphibian Species covered by this Checklist listed by listed by CITES EC- as well as Family Species Regulation EC 338/97 Regulation only 338/97 ANURA Aromobatidae Allobates femoralis X Aromobatidae Allobates hodli X Aromobatidae Allobates myersi X Aromobatidae Allobates zaparo X Aromobatidae Anomaloglossus rufulus X Bufonidae Altiphrynoides malcolmi X Bufonidae Altiphrynoides osgoodi X Bufonidae Amietophrynus channingi X Bufonidae Amietophrynus superciliaris X Bufonidae Atelopus zeteki X Bufonidae Incilius periglenes X Bufonidae Nectophrynoides asperginis X Bufonidae Nectophrynoides cryptus X Bufonidae Nectophrynoides frontierei X Bufonidae Nectophrynoides laevis X Bufonidae Nectophrynoides laticeps X Bufonidae Nectophrynoides minutus X Bufonidae Nectophrynoides paulae X Bufonidae Nectophrynoides poyntoni X Bufonidae Nectophrynoides pseudotornieri X Bufonidae Nectophrynoides tornieri X Bufonidae Nectophrynoides vestergaardi -

Reproductive Biology of Ameerega Trivittata(Anura: Dendrobatidae)
ACTA AMAZONICA http://dx.doi.org/10.1590/1809-4392201305384 Reproductive biology of Ameerega trivittata (Anura: Dendrobatidae) in an area of terra firme forest in eastern Amazonia Ellen Cristina Serrão ACIOLI1 * , Selvino NECKEL-OLIVEIRA2 ¹ Universidade Federal do Pará/Museu Paraense Emílio Goeldi, Programa de Pós Graduação em Zoologia, Av. Perimetral 1901/1907, Terra Firme, 66017-970, Belém, Pará, Brasil. ² Universidade Federal de Santa Catarina, Departamento de Ecologia e Zoologia, Centro de Ciências Biológicas, 88040-970, Florianópolis, Santa Catarina, Brasil. * Corresponding author: [email protected] ABSTRACT The reproductive success of tropical amphibians is influenced by factors such as body size and the characteristics of breeding sites. Data on reproductive biology are important for the understanding of population dynamics and the maintenance of species. The objectives of the present study were to examine the abundance ofAmeerega trivittata, analyze the use of microhabitats by calling males and the snout-vent length (SVL) of breeding males and females, the number of tadpoles carried by the males and mature oocytes in the females, as well as the relationship between the SVL of the female and both the number and mean size of the mature oocytes found in the ovaries. Three field trips were conducted between January and September, 2009. A total of 31 plots, with a mean area of 2.3 ha, were surveyed, resulting in records of 235 individuals, with a mean density of 3.26 individuals per hectare. Overall, 66.1% of the individuals sighted were located in the leaf litter, while 17.4% were perched on decaying tree trunks on the forest floor, 15.7% on the aerial roots ofCecropia trees, and 0.8% on lianas. -

Diet Composition of Ameerega Picta
Bonn zoological Bulletin 68 (1): 93–96 ISSN 2190–7307 2019 · Landgref Filho P. et al. http://www.zoologicalbulletin.de https://doi.org/10.20363/BZB-2019.68.1.093 Scientific note urn:lsid:zoobank.org:pub:CBA156F4-E9AA-401C-B036-77C362CE1E89 Diet composition of Ameerega picta (Tschudi, 1838) from the Serra da Bodoquena region in central Brazil, with a summary of dietary studies on species of the genus Ameerega (Anura: Dendrobatidae) Paulo Landgref Filho1, Fabrício H. Oda2, *, Fabio T. Mise3, Domingos de J. Rodrigues4 & Masao Uetanabaro5 1 Universidade Federal de Mato Grosso do Sul, Campus Aquidauana, 79200-000, Aquidauana, Mato Grosso do Sul, Brazil 2 Departamento de Química Biológica, Programa de Pós-graduação em Bioprospecção Molecular, Universidade Regional do Cariri, 63105-00, Crato, Ceará, Brazil 3 Departamento de Biologia, Universidade Estadual do Centro-Oeste, 85040-080, Guarapuava, Paraná, Brazil 4 Instituto de Ciências Naturais, Humanas e Sociais, Universidade Federal de Mato Grosso – Campus Universitário de Sinop, 78557-267, Sinop, Mato Grosso, Brazil 4 Instituto Nacional de Ciência e Tecnologia de Estudos Integrados da Biodiversidade Amazônica – Núcleo Regional de Sinop, Sinop, Mato Grosso, Brazil 5 Rua Clóvis n. 24, 79022-071, Campo Grande, Mato Grosso do Sul, Brazil * Corresponding author: Email: [email protected] 1 urn:lsid:zoobank.org:author:5821C96D-4D9E-4E1D-BE9B-C231B77D1AF3 2 urn:lsid:zoobank.org:author:349712B5-E1B2-4059-A826-58F081DD3A4D 3 urn:lsid:zoobank.org:author:E996AF2A-6CB7-4B73-8DEF-AB7C5EEE1AA0 4 urn:lsid:zoobank.org:author:ACFBA432-3220-4910-AFF5-8FF85053A872 5 urn:lsid:zoobank.org:author:626F6AE3-7D90-4500-BFDC-4E6F4E01B378 Abstract. -

Species Diversity and Conservation Status Of
Florida International University FIU Digital Commons Department of Biological Sciences College of Arts, Sciences & Education 4-2009 Species Diversity and Conservation Status of Amphibians in Madre De Dios, Southern Peru Rudolf Von May Department of Biological Sciences, Florida International University Karen Siu-Ting Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos Jennifer M. Jacobs San Francisco State University; California Academy of Sciences Margarita Medina-Muller Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos Giuseppe Gagliardi Museo de Zoología de la Universidad Nacional de la Amazonía Peruana See next page for additional authors Follow this and additional works at: https://digitalcommons.fiu.edu/cas_bio Part of the Biology Commons Recommended Citation May, Rudolf Von; Siu-Ting, Karen; Jacobs, Jennifer M.; Medina-Muller, Margarita; Gagliardi, Giuseppe; Rodriguez, Lily O.; and Donnelly, Maureen A., "Species Diversity and Conservation Status of Amphibians in Madre De Dios, Southern Peru" (2009). Department of Biological Sciences. 164. https://digitalcommons.fiu.edu/cas_bio/164 This work is brought to you for free and open access by the College of Arts, Sciences & Education at FIU Digital Commons. It has been accepted for inclusion in Department of Biological Sciences by an authorized administrator of FIU Digital Commons. For more information, please contact [email protected]. Authors Rudolf Von May, Karen Siu-Ting, Jennifer M. Jacobs, Margarita Medina-Muller, Giuseppe Gagliardi, Lily O. Rodriguez, and Maureen A. Donnelly This article is available at FIU Digital Commons: https://digitalcommons.fiu.edu/cas_bio/164 Herpetological Conservation and Biology 4(1):14-29 Submitted: 18 December 2007; Accepted: 4 August 2008 SPECIES DIVERSITY AND CONSERVATION STATUS OF AMPHIBIANS IN MADRE DE DIOS, SOUTHERN PERU 1,2 3 4,5 RUDOLF VON MAY , KAREN SIU-TING , JENNIFER M. -

October/November 2005
October/November 2005 Welcome to the second issue of Dartfrog ‘News’ Where does the time go? It seems like only yesterday we were sending out the maiden issue of this newsletter. So what has happened in between? Well, we went to the Bocas Del Toro archipelago in northeast Panama and to say that it was a dart frog paradise would be quite an understatement – it literally was dart frog heaven. In this issue we have the first instalment of our Bocas diary. We also have some photos of new dart frog acquisitions, of various tadpoles and a review of the excellent tree fern root or xaxim products. On the caudata front there are photos of egg/larvae development in the emperor salamander (Tylototriton shanjiing). As always there are other offers, some amazing frogs for sale that will not appear on our general web pricelist and tips and hints... “Amphibian of the month” Zaparo’s poison frog (Allobates (Epipedobates) zaparo) We first came across this species at one of the Dutch frog day shows near Amsterdam in October 2003. The specimens we picked up were rather small (about 10mm) and non- descript, just beginning to show the prominent yellow flash marks. Having never seen this species in true life we had no idea at the time what they would turn out like. We need not have worried – the dorsum gradually become more and more granular and an increasingly deep red in coloration. In the wild this species hails from Eastern Ecuador, northern Peru and possibly into southern Colombia and are concentrated around the humid forests of the Rio Pastaza and Rio Napo river basins. -

The Global Amphibian Trade Flows Through Europe: the Need for Enforcing and Improving Legislation
Biodivers Conserv DOI 10.1007/s10531-016-1193-8 REVIEW PAPER The global amphibian trade flows through Europe: the need for enforcing and improving legislation 1 2 3,4 Mark Auliya • Jaime Garcı´a-Moreno • Benedikt R. Schmidt • 1 5 6 Dirk S. Schmeller • Marinus S. Hoogmoed • Matthew C. Fisher • 7 1 8 Frank Pasmans • Klaus Henle • David Bickford • An Martel7 Received: 15 December 2015 / Revised: 11 August 2016 / Accepted: 13 August 2016 Ó Springer Science+Business Media Dordrecht 2016 Abstract The global amphibian trade is suspected to have brought several species to the brink of extinction, and has led to the spread of amphibian pathogens. Moreover, inter- national trade is not regulated for *98 % of species. Here we outline patterns and com- plexity underlying global amphibian trade, highlighting some loopholes that need to be addressed, focusing on the European Union. In spite of being one of the leading amphibian importers, the EU’s current legislation is insufficient to prevent overharvesting of those species in demand or the introduction and/or spread of amphibian pathogens into captive and wild populations. We suggest steps to improve the policy (implementation and enforcement) framework, including (i) an identifier specifically for amphibians in the Communicated by David Hawksworth. This article belongs to the Topical Collection: Biodiversity legal instruments and regulations. & Mark Auliya [email protected] & Jaime Garcı´a-Moreno [email protected] 1 Department of Conservation Biology, UFZ—Helmholtz Centre for Environmental Research,