Trna Biology Charges to the Front
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Inducible Gene Expression: Diverse Regulatory Mechanisms
REVIEWS MODES OF TRANSCRIPTIONAL REGULATION Inducible gene expression: diverse regulatory mechanisms Vikki M. Weake and Jerry L. Workman Abstract | The rapid activation of gene expression in response to stimuli occurs largely through the regulation of RNA polymerase II‑dependent transcription. In this Review, we discuss events that occur during the transcription cycle in eukaryotes that are important for the rapid and specific activation of gene expression in response to external stimuli. In addition to regulated recruitment of the transcription machinery to the promoter, it has now been shown that control steps can include chromatin remodelling and the release of paused polymerase. Recent work suggests that some components of signal transduction cascades also play an integral part in activating transcription at target genes. Chromatin Cells must be able to rapidly respond to changes in RNA polymerase II (Pol II)‑dependent transcription. A nucleoprotein structure their external environment — such as temperature or Although a traditional model of activator‑dependent formed of repeating nutrient availability — to exploit and survive in new recruitment of Pol II and the general transcription nucleosomal units in which conditions. Even cells in a multicellular organism need factors (GTFs) holds true for many inducible genes, 147 base pairs of DNA are wrapped around an octamer of to respond to developmental cues such as signalling recent studies suggest that Pol II is already present and 1–6 histone proteins consisting molecules to determine when to divide, migrate or poised for transcription at many inducible genes . of an H3–H4 tetramer flanked die. The production of new proteins in response to Therefore, it is becoming increasingly apparent that by two H2A–H2B dimers external stimuli results largely from rapid activation there is an additional level of regulation that occurs of gene transcription — this is known as inducible during the initial stages of transcription elongation Co-activator A protein that is recruited to gene expression. -

A Genome-Wide Screen for Genes Affecting Spontaneous Direct-Repeat Recombination In
bioRxiv preprint doi: https://doi.org/10.1101/2020.02.11.943795; this version posted February 11, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 A genome-wide screen for genes affecting spontaneous direct-repeat recombination in 2 Saccharomyces cerevisiae 3 4 5 Daniele Novarina*, Ridhdhi Desai†, Jessica A. Vaisica†, Jiongwen Ou†, Mohammed Bellaoui†,1, 6 Grant W. Brown†,2 and Michael Chang*,3 7 8 *European Research Institute for the Biology of Ageing, University of Groningen, University 9 Medical Center Groningen, 9713 AV Groningen, the Netherlands 10 †Department of Biochemistry and Donnelly Centre, University of Toronto, Toronto, ON M5S 11 3E1, Canada 12 13 1Current address: Genetics Unit, Faculty of Medicine and Pharmacy, University Mohammed 14 Premier, Oujda, Morocco 15 16 2Co-corresponding author: Department of Biochemistry and Donnelly Centre, University of 17 Toronto, 160 College Street, Toronto, ON M5S 3E1 Canada. E-mail: [email protected] 18 3Co-corresponding author: European Research Institute for the Biology of Ageing, University of 19 Groningen, University Medical Center Groningen, Antonius Deusinglaan 1, 9713 AV Groningen, 20 the Netherlands. E-mail: [email protected] 21 1 bioRxiv preprint doi: https://doi.org/10.1101/2020.02.11.943795; this version posted February 11, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. -

Yeast Genome Gazetteer P35-65
gazetteer Metabolism 35 tRNA modification mitochondrial transport amino-acid metabolism other tRNA-transcription activities vesicular transport (Golgi network, etc.) nitrogen and sulphur metabolism mRNA synthesis peroxisomal transport nucleotide metabolism mRNA processing (splicing) vacuolar transport phosphate metabolism mRNA processing (5’-end, 3’-end processing extracellular transport carbohydrate metabolism and mRNA degradation) cellular import lipid, fatty-acid and sterol metabolism other mRNA-transcription activities other intracellular-transport activities biosynthesis of vitamins, cofactors and RNA transport prosthetic groups other transcription activities Cellular organization and biogenesis 54 ionic homeostasis organization and biogenesis of cell wall and Protein synthesis 48 plasma membrane Energy 40 ribosomal proteins organization and biogenesis of glycolysis translation (initiation,elongation and cytoskeleton gluconeogenesis termination) organization and biogenesis of endoplasmic pentose-phosphate pathway translational control reticulum and Golgi tricarboxylic-acid pathway tRNA synthetases organization and biogenesis of chromosome respiration other protein-synthesis activities structure fermentation mitochondrial organization and biogenesis metabolism of energy reserves (glycogen Protein destination 49 peroxisomal organization and biogenesis and trehalose) protein folding and stabilization endosomal organization and biogenesis other energy-generation activities protein targeting, sorting and translocation vacuolar and lysosomal -
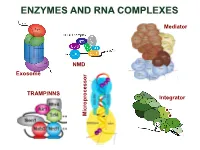
Enzymes and Rna Complexes
ENZYMES AND RNA COMPLEXES Mediator NMD Exosome NMD TRAMP/NNS Integrator Microprocessor RNA PROCESSING and DECAY machinery: RNases Protein Function Characteristics Exonucleases 5’ 3’ Xrn1 cytoplasmic, mRNA degradation processsive Rat1 nuclear, pre-rRNA, sn/snoRNA, pre-mRNA processing and degradation Rrp17/hNol12 nuclear, pre-rRNA processing Exosome 3’ 5’ multisubunit exo/endo complex subunits organized as in bacterial PNPase Rrp44/Dis3 catalytic subunit Exo/PIN domains, processsive Rrp4, Rrp40 pre-rRNA, sn/snoRNA processing, mRNA degradation Rrp41-43, 45-46 participates in NMD, ARE-dependent, non-stop decay Mtr3, Ski4 Mtr4 nuclear helicase cofactor DEAD box Rrp6 (Rrp47) nuclear exonuclease ( Rrp6 BP, cofactor) RNAse D homolog, processsive Ski2,3,7,8 cytoplasmic exosome cofactors. SKI complex helicase, GTPase Other 3’ 5’ Rex1-4 3’-5’ exonucleases, rRNA, snoRNA, tRNA processing RNase D homolog DXO 3’-5’ exonuclease in addition to decapping mtEXO 3’ 5’ mitochondrial degradosome RNA degradation in yeast Suv3/ Dss1 helicase/ 3’-5’ exonuclease DExH box/ RNase II homolog Deadenylation Ccr4/NOT/Pop2 major deadenylase complex (Ccr, Caf, Pop, Not proteins) Ccr4- Mg2+ dependent endonuclease Pan2p/Pan3 additional deadenylases (poliA tail length) RNase D homolog, poly(A) specific nuclease PARN mammalian deadenylase RNase D homolog, poly(A) specific nuclease Endonucleases RNase III -Rnt1 pre-rRNA, sn/snoRNA processing, mRNA degradation dsRNA specific -Dicer, Drosha siRNA/miRNA biogenesis, functions in RNAi PAZ, RNA BD, RNase III domains Ago2 Slicer -

A Disease-Linked Lncrna Mutation in Rnase MRP Inhibits Ribosome Synthesis
bioRxiv preprint doi: https://doi.org/10.1101/2021.03.29.437572; this version posted March 29, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. A disease-linked lncRNA mutation in RNase MRP inhibits ribosome synthesis Nic Roberston1, Vadim Shchepachev1, David Wright2, Tomasz W. Turowski1, Christos Spanos1, Aleksandra Helwak1, Rose Zamoyska2, David Tollervey1 1 Wellcome Centre for Cell Biology, University of Edinburgh, Edinburgh, UK 2 Ashworth Laboratories, Institute of Immunology and Infection Research, University of Edinburgh, Edinburgh, UK Keywords: protein-RNA interaction; RNA-binding sites; UV crosslinking; mass spectrometry; genetic disease; Cartilage Hair Hypoplasia; ribosome synthesis; T cell activation Running title: RNase MRP defects cause ribosomopathy Highlights: • Mutations in RMRP lncRNA impair pre-rRNA processing and T cell activation • Patient derived fibroblasts show impaired pre-rRNA processing • Cells with the most common disease-linked mutation have specific processing defects • Cytoplasmic ribosomes and intact RNase MRP complexes are also reduced in these cells 1 bioRxiv preprint doi: https://doi.org/10.1101/2021.03.29.437572; this version posted March 29, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. Abstract Mutations in the human RMRP gene cause Cartilage Hair Hypoplasia (CHH), an autosomal recessive disorder characterized by skeletal abnormalities and impaired T cell activation. -

Genetic Analysis of the Hsm3 Protein Function in Yeast Saccharomyces Cerevisiae Nub4 Complex
G C A T T A C G G C A T genes Article Genetic Analysis of the Hsm3 Protein Function in Yeast Saccharomyces cerevisiae NuB4 Complex Tatiyana A. Evstyukhina 1,2, Elena A. Alekseeva 1,2,* , Dmitriy V. Fedorov 1, Vyacheslav T. Peshekhonov 1,2 and Vladimir G. Korolev 1,2 1 Laboratory of Eukaryotic Genetics, Department of Molecular and Radiation Biophysics, Petersburg Nuclear Physics Institute Named by B.P. Konstantinov of National Research Centre “Kurchatov Institute”, 188300 Gatchina, Russia; [email protected] (T.A.E.); [email protected] (D.V.F.); [email protected] (V.T.P.); [email protected] (V.G.K.) 2 Laboratory of Molecular Genetic and Recombination Technologies, Kurchatov Genome Center— Petersburg Nuclear Physics Institute, mkr. Orlova Roscha 1, Leningrad District, 188300 Gatchina, Russia * Correspondence: [email protected] Abstract: In the nuclear compartment of yeast, NuB4 core complex consists of three proteins, Hat1, Hat2, and Hif1, and interacts with a number of other factors. In particular, it was shown that NuB4 complex physically interacts with Hsm3p. Early we demonstrated that the gene HSM3 participates in the control of replicative and reparative spontaneous mutagenesis, and that hsm3D mutants increase the frequency of mutations induced by different mutagens. It was previously believed that the HSM3 gene controlled only some minor repair processes in the cell, but later it was suggested that it had a chaperone function with its participation in proteasome assembly. In this work, we analyzed the properties of three hsm3D, hif1D, and hat1D mutants. The results obtained showed that the Citation: Evstyukhina, T.A.; Hsm3 protein may be a functional subunit of NuB4 complex. -

A Specialized Processing Body That Is Temporally and Asymmetrically Regulated During the Cell Cycle in Saccharomyces Cerevisiae
JCB: ARTICLE A specialized processing body that is temporally and asymmetrically regulated during the cell cycle in Saccharomyces cerevisiae Tina Gill, Jason Aulds, and Mark E. Schmitt Department of Biochemistry and Molecular Biology, State University of New York Upstate Medical University, Syracuse, NY 13210 Nase mitochondrial RNA processing (MRP) is an this spot was asymmetrically found in daughter cells, essential ribonucleoprotein endoribonuclease that where the RNase MRP substrate, CLB2 mRNA, localizes. R functions in the degradation of specifi c mRNAs in- Both the mitotic exit network and fourteen early ana- volved in cell cycle regulation. We have investigated phase release pathways are nonessential but important where this processing event occurs and how it is regu- for the temporal changes in localization. Asymmetric lo- lated. As expected, results demonstrate that RNase MRP calization was found to be dependent on the locasome. is predominantly localized in the nucleolus, where it The evidence suggests that these spots are specialized processes ribosomal RNAs. However, after the initiation processing bodies for the degradation of transcripts that of mitosis, RNase MRP localizes throughout the entire are cell cycle regulated and daughter cell localized. We nucleus and in a single discrete cytoplasmic spot that have called these TAM bodies for temporal asymmetric persists until the completion of telophase. Furthermore, MRP bodies. Introduction RNase mitochondrial RNA processing (MRP) is an essential ri- 27SA preribosomal RNA at the A3 site, forming the 5.8S(s) bonucleoprotein endoribonuclease that cleaves RNA substrates ribosomal RNA (rRNA; Schmitt and Clayton, 1993; Lygerou in a site-specifi c manner and is highly conserved in eukaryotes et al., 1996). -

Functional Link Between Mitochondria and Rnr3, the Minor Catalytic Subunit of Yeast Ribonucleotide Reductase
Research Article www.microbialcell.com Functional link between mitochondria and Rnr3, the minor catalytic subunit of yeast ribonucleotide reductase Isaac Corcoles-Saez1, Jean-Luc Ferat2, Michael Costanzo3, Charles M. Boone3 and Rita S. Cha1,* 1 School of Medical Sciences, North West Cancer Research Institute, Bangor University, Deniol Road, Bangor, LL57 2UW, United Kingdom. 2 Institute of Integrative Biology of the Cell (I2BC), Avenue de la Terrasse, Paris, France. 3 University of Toronto, Donnelly Centre, 160 College Street, Toronto, Ontario, M5S 3E1, Canada.. * Corresponding Author: Rita S. Cha, Tel: +44 (0)1248 38286; E-mail: [email protected] ABSTRACT Ribonucleotide reductase (RNR) is an essential holoenzyme re- doi: 10.15698/mic2019.06.680 quired for de novo synthesis of dNTPs. The Saccharomyces cerevisiae genome Received originally: 24.12.2019; encodes for two catalytic subunits, Rnr1 and Rnr3. While Rnr1 is required for in revised form: 09.05.2019, DNA replication and DNA damage repair, the function(s) of Rnr3 is unknown. Accepted 13.05.2019, Published 20.05.2019. Here, we show that carbon source, an essential nutrient, impacts Rnr1 and Rnr3 abundance: Non-fermentable carbon sources or limiting concentrations of glucose down regulate Rnr1 and induce Rnr3 expression. Oppositely, abun- Keywords: Rnr1, Rnr3, Mec1, carbon dant glucose induces Rnr1 expression and down regulates Rnr3. The carbon source, respiration, mitochondria, source dependent regulation of Rnr3 is mediated by Mec1, the budding yeast dNTP. ATM/ATR checkpoint response kinase. Unexpectedly, this regulation is inde- pendent of all currently known components of the Mec1 DNA damage re- Abbreviations: sponse network, including Rad53, Dun1, and Tel1, implicating a novel Mec1 DDR – DNA damage response, signalling axis. -

Ribonuclease A
Chem. Rev. 1998, 98, 1045−1065 1045 Ribonuclease A Ronald T. Raines Departments of Biochemistry and Chemistry, University of WisconsinsMadison, Madison, Wisconsin 53706 Received October 10, 1997 (Revised Manuscript Received January 12, 1998) Contents I. Introduction 1045 II. Heterologous Production 1046 III. Structure 1046 IV. Folding and Stability 1047 A. Disulfide Bond Formation 1047 B. Prolyl Peptide Bond Isomerization 1048 V. RNA Binding 1048 A. Subsites 1048 B. Substrate Specificity 1049 C. One-Dimensional Diffusion 1049 D. Processive Catalysis 1050 VI. Substrates 1050 VII. Inhibitors 1051 Ronald T. Raines was born in 1958 in Montclair, NJ. He received Sc.B. VIII. Reaction Mechanism 1052 degrees in chemistry and biology from the Massachusetts Institute of A. His12 and His119 1053 Technology. At M.I.T., he worked with Christopher T. Walsh to reveal the reaction mechanisms of pyridoxal 5′-phosphate-dependent enzymes. B. Lys41 1054 Raines was a National Institutes of Health predoctoral fellow in the C. Asp121 1055 chemistry department at Harvard University. There, he worked with D. Gln11 1056 Jeremy R. Knowles to elucidate the reaction energetics of triosephosphate IX. Reaction Energetics 1056 isomerase. Raines was a Helen Hay Whitney postdoctoral fellow in the biochemistry and biophysics department at the University of California, A. Transphosphorylation versus Hydrolysis 1056 San Francisco. At U.C.S.F., he worked with William J. Rutter to clone, B. Rate Enhancement 1057 express, and mutate the cDNA that codes for ribonuclease A. Raines X. Ribonuclease S 1058 then joined the faculty of the biochemistry department at the University s A. S-Protein−S-Peptide Interaction 1058 of Wisconin Madison, where he is now associate professor of biochem- istry and chemistry. -

Characterization of the Mammalian RNA Exonuclease 5/NEF-Sp As a Testis-Specific Nuclear 3′′′′′ → 5′′′′′ Exoribonuclease
Downloaded from rnajournal.cshlp.org on October 7, 2021 - Published by Cold Spring Harbor Laboratory Press Characterization of the mammalian RNA exonuclease 5/NEF-sp as a testis-specific nuclear 3′′′′′ → 5′′′′′ exoribonuclease SARA SILVA,1,2 DAVID HOMOLKA,1 and RAMESH S. PILLAI1 1Department of Molecular Biology, University of Geneva, CH-1211 Geneva 4, Switzerland 2European Molecular Biology Laboratory, Grenoble Outstation, 38042, France ABSTRACT Ribonucleases catalyze maturation of functional RNAs or mediate degradation of cellular transcripts, activities that are critical for gene expression control. Here we identify a previously uncharacterized mammalian nuclease family member NEF-sp (RNA exonuclease 5 [REXO5] or LOC81691) as a testis-specific factor. Recombinant human NEF-sp demonstrates a divalent metal ion-dependent 3′′′′′ → 5′′′′′ exoribonuclease activity. This activity is specific to single-stranded RNA substrates and is independent of their length. The presence of a 2′′′′′-O-methyl modification at the 3′′′′′ end of the RNA substrate is inhibitory. Ectopically expressed NEF-sp localizes to the nucleolar/nuclear compartment in mammalian cell cultures and this is dependent on an amino-terminal nuclear localization signal. Finally, mice lacking NEF-sp are viable and display normal fertility, likely indicating overlapping functions with other nucleases. Taken together, our study provides the first biochemical and genetic exploration of the role of the NEF-sp exoribonuclease in the mammalian genome. Keywords: NEF-sp; LOC81691; Q96IC2; REXON; RNA exonuclease 5; REXO5; 2610020H08Rik INTRODUCTION clease-mediated processing to create their final 3′ ends: poly(A) tails of most mRNAs or the hairpin structure of Spermatogenesis is the process by which sperm cells are replication-dependent histone mRNAs (Colgan and Manley produced in the male germline. -

Protein Engineering to Exploit and Explore Bovine Secretory Ribonucleases
PROTEIN ENGINEERING TO EXPLOIT A N D EXPLORE BOVINE SECRETORY RIBONUCLEASES by JIN-SOO KIM A thesis submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Biochemistry) at the UNIVERSITY OF WISCONSIN-MADISON 1994 Reproduced with permission of the copyright owner. Further reproduction prohibited without permission. ACKNOWLEDGEMENTS I would like to thank Dr. Ronald T. Raines for his advice and support. His scientific insight has been very helpful throughout this work. I would also like to thank the entire Raines group for their friendship and companionship. I am grateful to Dr. J. Soucek and Dr. J. Matousek for their collaboration with us, which has been a valuable part of the BS-RNase research. I thank Dr. M. Karpeisky for suggesting the protein fusion project, and Dr. G. D'Alessio and Dr. L. Mazzarella for providing the coordinates of BS-RNase. I have been generously supported by Steenbock predoctoral fellowship from the Department of Biochemistry. Finally, I thank my parents, who have encouraged (or at least not discouraged) me to pursue a career in science since I was a kid. Reproduced with permission of the copyright owner. Further reproduction prohibited without permission ABSTRACT PROTEIN ENGINEERING TO EXPLOIT AND EXPLORE BOVINE SECRETORY RIBONUCLEASES Jin-Soo Kim Under the supervision of Dr. Ronald T. Raines at the University of Wisconsin-Madison Ribonuclease S-peptide (residues 1-20) and S-protein (residues 21- 124) are the enzymatically inactive products of the limited digestion of bovine pancreatic ribonuclease A (RNase A) by subtilisin. S-Peptide binds S-protein with high affinity to form RNase S, which has full enzymatic activity. -

Generate Metabolic Map Poster
Authors: Pallavi Subhraveti Ron Caspi Peter Midford Peter D Karp An online version of this diagram is available at BioCyc.org. Biosynthetic pathways are positioned in the left of the cytoplasm, degradative pathways on the right, and reactions not assigned to any pathway are in the far right of the cytoplasm. Transporters and membrane proteins are shown on the membrane. Ingrid Keseler Periplasmic (where appropriate) and extracellular reactions and proteins may also be shown. Pathways are colored according to their cellular function. Gcf_003855395Cyc: Shewanella livingstonensis LMG 19866 Cellular Overview Connections between pathways are omitted for legibility.