Secretion by Fibronectin and Type I Collagen of Human Airway Smooth
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Collagen and Elastin Fibres
J Clin Pathol: first published as 10.1136/jcp.s3-12.1.49 on 1 January 1978. Downloaded from J. clin. Path., 31, Suppl. (Roy. Coll. Path.), 12, 49-58 Collagen and elastin fibres A. J. BAILEY From the Agricultural Research Council, Meat Research Institute, Langford, Bristol Although an understanding of the intracellular native collagen was generated from type I pro- biosynthesis of both collagen and elastin is of collagen. Whether this means that the two pro- considerable importance it is the subsequent extra- collagens are converted by different enzyme systems cellular changes involving fibrogenesis and cross- and the type III enzyme was deficient in these linking that ensure that these proteins ultimately fibroblast cultures, or that the processing of pro become the major supporting tissues of the body. type III is extremely slow, is not known. The latter This paper summarises the formation and stability proposal is consistent with the higher proportion of collagen and elastin fibres. of soluble pro type III extractable from tissue (Lenaers and Lapiere, 1975; Timpl et al., 1975). Collagen Basement membrane collagens, on the other hand, do not form fibres and this property may be The non-helical regions at the ends of the triple due to the retention of the non-helical extension helix of procollagen probably provide a number of peptides (Kefalides, 1973). In-vivo biosynthetic different intracellular functions-that is, initiating studies showing the absence of any extension peptide rapid formation of the triple helix; inhibiting intra- removal support this (Minor et al., 1976), but other cellular fibrillogenesis; and facilitating transmem- workers have reported that there is some cleavage brane movement. -

Evaluation of Elastin/Collagen Content in Human Dermis In-Vivo by Multiphoton Tomography—Variation with Depth and Correlation with Aging
Cosmetics 2014, 1, 211-221; doi:10.3390/cosmetics1030211 OPEN ACCESS cosmetics ISSN 2079-9284 www.mdpi.com/journal/cosmetics Article Evaluation of Elastin/Collagen Content in Human Dermis in-Vivo by Multiphoton Tomography—Variation with Depth and Correlation with Aging Jean-Christophe Pittet 1,*, Olga Freis 2,†, Marie-Danielle Vazquez-Duchêne 2,†, Gilles Périé 2,† and Gilles Pauly 2,† 1 Orion Concept, 100 Rue de Suède, 37100 Tours, France 2 BASF Beauty Care Solutions France SAS, 3 Rue de Seichamps, CS 71040 Pulnoy, 54272 Essey-lès-Nancy Cedex, France; E-Mails: [email protected] (O.F.); [email protected] (M.-D.V.-D.); [email protected] (G.Pé.); [email protected] (G.Pa.) † These authors contributed equally to this work. * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +33-247-052-316; Fax: +33-610-786-695. Received: 14 March 2014; in revised form: 31 July 2014 / Accepted: 1 August 2014 / Published: 20 August 2014 Abstract: The aim of this study was to evaluate the influence of the depth of the dermis on the measured collagen and elastin levels and to establish the correlation between the amount of these two extracellular matrix (ECM) components and age. Multiphoton Microscopy (MPM) that measures the autofluorescence (AF) and second harmonic generation (SHG) was used to quantify the levels of elastin and collagen and to determine the SAAID (SHG-to-AF Aging Index of Dermis) at two different skin depths. A 50 MHz ultrasound scanner was used for the calculation of the Sub Epidermal Non Echogenic Band (SENEB). -

Blood Vitronectin Is a Major Activator of LIF and IL-6 in the Brain Through Integrin–FAK and Upar Signaling Matthew P
© 2018. Published by The Company of Biologists Ltd | Journal of Cell Science (2018) 131, jcs202580. doi:10.1242/jcs.202580 RESEARCH ARTICLE Blood vitronectin is a major activator of LIF and IL-6 in the brain through integrin–FAK and uPAR signaling Matthew P. Keasey1, Cuihong Jia1, Lylyan F. Pimentel1,2, Richard R. Sante1, Chiharu Lovins1 and Theo Hagg1,* ABSTRACT Microglia and astrocytes express the VTN receptors αvβ3 and α β We defined how blood-derived vitronectin (VTN) rapidly and potently v 5 integrin (Herrera-Molina et al., 2012; Kang et al., 2008; activates leukemia inhibitory factor (LIF) and pro-inflammatory Milner, 2009; Welser-Alves et al., 2011). Microglia and astrocytes, interleukin 6 (IL-6) in vitro and after vascular injury in the brain. as well as endothelial cells, are major producers of pro- α in vitro Treatment with VTN (but not fibrinogen, fibronectin, laminin-111 or inflammatory cytokines, such as IL-6 and TNF , and collagen-I) substantially increased LIF and IL-6 within 4 h in after traumatic or ischemic injury to the brain (Banner et al., 1997; C6-astroglioma cells, while VTN−/− mouse plasma was less effective Erta et al., 2012; Lau and Yu, 2001) or upon self-induction by IL-6 than that from wild-type mice. LIF and IL-6 were induced by (Van Wagoner and Benveniste, 1999). IL-6 is a major regulator of a intracerebral injection of recombinant human (rh)VTN in mice, but variety of inflammatory disorders and a target for therapies (Hunter induction seen upon intracerebral hemorrhage was less in VTN−/− and Jones, 2015). -

With Caviar, Keratin & Collagen
WITH CAVIAR, KERATIN & COLLAGEN Professional treatments for colour, bleaching, care and maintenance with CAVIAR, KERATIN and COLLAGEN Professional styling products with CAVIAR, KERATIN and COLLAGEN Technical professional products with CAVIAR, KERATIN and COLLAGEN 2 Professional products by Very high technology professional products to colour, treat and protect hair from the continuous chemical and environmental stress caused on a daily basis. Formulas based on: Caviar Keratin Collagen 3 WITH CAVIAR, KERATIN & COLLAGEN Colouring permanentCream professional ammonia PPD • Respects the hair structure thanks to an exposure free free time of 12 minutes • Non-progressive • Ammonia free • Paraphenylenediamine free • Unleashes all the EFFICANCY of its active principles and maximum COLOURING POWER Mix 1 : 1 4 WITH CAVIAR, KERATIN & COLLAGEN 1. Respect for scalp and hair thanks to a shorter processing time 2. Maximum grey hair coverage 3. Lightens up to 4 tones 4. Ammonia free and Paraphenylenediamine free 5. Safe application even on customers with a sensitive scalp 6. Extreme colour brilliancy and uniformity 7. Very easy and practical to use 8. Long lasting reflections 9. High colour fastness 10. Great protection action 11. Effective restructuring action benefits 12. Maximum conditioning 5 WITH CAVIAR, KERATIN & COLLAGEN Benefits 1 2 3 Respect for scalp and hair thanks to a shorter processing Lightens up to Maximum grey hair 4 tones time coverage 6 WITH CAVIAR, KERATIN & COLLAGEN has been developed according Cosmetic colour pDT BASE Be Colour 12 Minute Dpe DIAMINOPHENOXYETHANOL to the rules of the “molar stoichiometry,” a technique creamy gel with RESORCINOL m-AMINOPHENOL of colouring clear and uncompromising. It is based on CAVIAR, a mathematical principle according to which the molar concentration of the dye base is equal to the molar KERATIN and concentration of the sum of the other colouring couplers. -
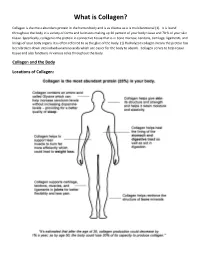
What Is Collagen?
What is Collagen? Collagen is the most abundant protein in the human body and is as diverse as it is multifunctional (1). It is found throughout the body in a variety of forms and functions making up 30 percent of your body tissue and 70 % of your skin tissue. Specifically, collagen is the protein in connective tissue that is in bone marrow, tendons, cartilage, ligaments, and linings of your body organs. It is often referred to as the glue of the body. (1) Hydrolyzed collagen means the protein has been broken down into individual amino acids which are easier for the body to absorb. Collagen serves to help repair tissue and also functions in various roles throughout the body. Collagen and the Body Locations of Collagen: What Does Collagen Do? Collagen has numerous structural properties but also plays a vital role in the repair of almost all the body’s tissues. Some diseases are directly linked to lacking this essential protein. Depending on which part of the body it is located, collagen serves different purposes. In skin: Found in the inner layer, this connective tissue gives the skin its structure and strength and also functions in the replacement of dead skin cells. A lack of collagen in the skin can contribute to a decrease in skin health leading to stretch marks, dark spots, and infections as well as affecting the skin’s ability to maintain moisture. In internal organs and blood vessels: In the lining of your organs like in the stomach, kidneys, blood vessels and spleen, collagen functions as a protective covering and a fibrous barrier. -

Characterization of Collagen Fibers (I, III, IV) and Elastin of Normal and Neoplastic Canine Prostatic Tissues
veterinary sciences Article Characterization of Collagen Fibers (I, III, IV) and Elastin of Normal and Neoplastic Canine Prostatic Tissues Luis Gabriel Rivera Calderón 1, Priscila Emiko Kobayashi 2, Rosemeri Oliveira Vasconcelos 1, Carlos Eduardo Fonseca-Alves 3 and Renée Laufer-Amorim 2,* 1 Department of Veterinary Pathology, School of Agricultural and Veterinarian Sciences, São Paulo State University (Unesp), Jaboticabal, São Paulo 14884-900, Brazil; [email protected] (L.G.R.C.); [email protected] (R.O.V.) 2 Department of Veterinary Clinic, School of Veterinary Medicine and Animal Science, São Paulo State University (Unesp), Botucatu, São Paulo 18618-681, Brazil; [email protected] 3 Department of Veterinary Surgery and Anesthesiology, School of Veterinary Medicine and Animal Science, São Paulo State University (Unesp), Botucatu, São Paulo 18618-681, Brazil; [email protected] * Correspondence: [email protected]; Tel.: +55-14-3880-2076 Received: 7 January 2019; Accepted: 25 February 2019; Published: 2 March 2019 Abstract: This study aimed to investigate collagen (Coll-I, III, IV) and elastin in canine normal prostate and prostate cancer (PC) using Picrosirius red (PSR) and Immunohistochemical (IHC) analysis. Eight normal prostates and 10 PC from formalin-fixed, paraffin-embedded samples were used. Collagen fibers area was analyzed with ImageJ software. The distribution of Coll-I and Coll-III was approximately 80% around prostatic ducts and acini, 15% among smooth muscle, and 5% surrounding blood vessels, in both normal prostate and PC. There was a higher median area of Coll-III in PC when compared to normal prostatic tissue (p = 0.001 for PSR and p = 0.05 for IHC). -

Empowering Collagen Targeting for Diagnosis and Treatment of Human Conditions
EmpoweringEmpowering collagen collagen targetingtargeting for for diagnosis prognosis of fibrotic and treatment of Manka,SW. 2012 conditions.human conditions. 1 3Helix as a platform diagnostic company 2020 2022 2030 Innovative research reagent for Clinic histopathology providing Platform fibrotic prognostic detection of collagen damage best in class prognostic ability in within multi-disease states liver fibrosis (NAFLD, NASH) Strengthening IP portfolio with market Clinical histopathology AND Non-Invasive approaches serum testing and medical disrupting products while subsequently Analytic specific reagent to allow for fast imaging developing strong partnerships with world market access leading companies for commercialization Targeting impactful markets of IPF, kidney Focused on paired biopsy research and fibrosis, AMD, Keloids and cardiac fibrosis collaboration with clinical laboratories. in addition to fibrotic liver diseases. 2 Liver Fibrosis market is GROWING Fatty Liver Fibrotic Liver Healthy Liver Cirrhosis NAFLD NASH USA Epidemiology 328 Million 83 Million 16 Million 1.5 Million (2015) Predicted USA Epidemiology 360 Million 101 Million 27 Million 3.4 Million (2030) Estes,C. 2018 3 NASH Therapeutics are finishing clinical trials and are coming to market. • VK2809 Phase II • OCALIVA (OCA) • NDA Filed • $78,000/year current cost • $20,000 predicted • Resmetirom Phase III • 23% respond to treatment 16 Million NASH patients in the USA * $20,000 • $320 Billion Annual Cost for treatable market Aramchol Phase III/IV 4 Stratification of patient population is needed to reduce unnecessary therapeutic intervention. • Progression of NAFL and 100% NASH is variable patient to NAFLD patient. 33% • Prediction of the progression can modify the Fibrosis Progression 20% disease intervention and treatment. Rapid Fibrosis • No product on the market Progression (stage 0 to stage today is equipped for 3/4 over 5.9 years) prognosis of liver fibrosis Singh, S. -

Collagen Type I and Decorin Expression in Tenocytes Depend On
Wagenhäuser et al. BMC Musculoskeletal Disorders 2012, 13:140 http://www.biomedcentral.com/1471-2474/13/140 RESEARCH ARTICLE Open Access Collagen type I and decorin expression in tenocytes depend on the cell isolation method Markus U Wagenhäuser1†, Matthias F Pietschmann1*†, Birte Sievers1, Denitsa Docheva2, Matthias Schieker2, Volkmar Jansson1 and Peter E Müller1 Abstract Backround: The treatment of rotator cuff tears is still challenging. Tendon tissue engineering (TTE) might be an alternative in future. Tenocytes seem to be the most suitable cell type as they are easy to obtain and no differentiation in vitro is necessary. The aim of this study was to examine, if the long head of the biceps tendon (LHB) can deliver viable tenocytes for TTE. In this context, different isolation methods, such as enzymatic digestion (ED) and cell migration (CM), are investigated on differences in gene expression and cell morphology. Methods: Samples of the LHB were obtained from patients, who underwent surgery for primary shoulder arthroplasty. Using ED as isolation method, 0.2% collagenase I solution was used. Using CM as isolation method, small pieces of minced tendon were put into petri-dishes. After cell cultivation, RT-PCR was performed for collagen type I, collagen type III, decorin, tenascin-C, fibronectin, Scleraxis, tenomodulin, osteopontin and agreccan. Results: The total number of isolated cells, in relation to 1 g of native tissue, was 14 times higher using ED. The time interval for cell isolation was about 17 hours using ED and approximately 50 days using CM. Cell morphology in vitro was similar for both isolation techniques. Higher expression of collagen type I could be observed in tenocyte-like cell cultures (TLCC) using ED as isolation method (p < 0.05), however decorin expression was higher in TLCC using CM as isolation method (p < 0.05). -

A Comparison of in Vivo Gene Delivery Methods for Antisense Therapy in Ligament Healing
Gene Therapy (1998) 5, 1455–1461 1998 Stockton Press All rights reserved 0969-7128/98 $12.00 http://www.stockton-press.co.uk/gt A comparison of in vivo gene delivery methods for antisense therapy in ligament healing N Nakamura1, SA Timmermann1, DA Hart1, Y Kaneda2, NG Shrive1, K Shino3, T Ochi3 and CB Frank1 1McCaig Centre for Joint Injury and Arthritis Research, University of Calgary, Calgary, Alberta, Canada; 2Institute for Molecular and Cellular Biology, Osaka University; and 3Department of Orthopaedic Surgery, Osaka University Medical School, Osaka, Japan To determine the most efficient in vivo delivery method of at 7 days after transfection. We then introduced antisense oligonucleotides for antisense therapy in ligament healing, ODN for the rabbit proteoglycan, decorin, into ligament fluorescence-labelled phosphorothioate oligodeoxynuleo- scars with this delivery method and confirmed a significant tides (ODN) were introduced into 12 rabbit ligament scars inhibition of decorin mRNA expression in antisense-treated 2 weeks after injury using haemagglutinating virus of Japan scar tissues in vivo both at 2 days (42.3 ± 14.7% of sense (Sendai virus; HVJ)-conjugated liposomes. We compared control ± s.d.; P Ͻ 0.0025) and 3 weeks (60.5 ± 28.2% of the efficiency of cellular uptake of fluorescence as a per- sense control ± s.d.; P Ͻ 0.024) after treatment, compared centage of all cells in each scar using three delivery pro- with sense ODN-treated scars. Decorin was significantly cedures: (1) direct free-hand injection into the ligament suppressed also at protein level in antisense-treated scars scar using a conventional syringe; (2) systematic direct at 4 weeks (66.6 ± 35.7% of sense control ± s.d.; scar injection using a repeating 10 l dispenser and a P Ͻ 0.045) after treatment. -

Collagens—Structure, Function, and Biosynthesis
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by University of East Anglia digital repository Advanced Drug Delivery Reviews 55 (2003) 1531–1546 www.elsevier.com/locate/addr Collagens—structure, function, and biosynthesis K. Gelsea,E.Po¨schlb, T. Aignera,* a Cartilage Research, Department of Pathology, University of Erlangen-Nu¨rnberg, Krankenhausstr. 8-10, D-91054 Erlangen, Germany b Department of Experimental Medicine I, University of Erlangen-Nu¨rnberg, 91054 Erlangen, Germany Received 20 January 2003; accepted 26 August 2003 Abstract The extracellular matrix represents a complex alloy of variable members of diverse protein families defining structural integrity and various physiological functions. The most abundant family is the collagens with more than 20 different collagen types identified so far. Collagens are centrally involved in the formation of fibrillar and microfibrillar networks of the extracellular matrix, basement membranes as well as other structures of the extracellular matrix. This review focuses on the distribution and function of various collagen types in different tissues. It introduces their basic structural subunits and points out major steps in the biosynthesis and supramolecular processing of fibrillar collagens as prototypical members of this protein family. A final outlook indicates the importance of different collagen types not only for the understanding of collagen-related diseases, but also as a basis for the therapeutical use of members of this protein family discussed in other chapters of this issue. D 2003 Elsevier B.V. All rights reserved. Keywords: Collagen; Extracellular matrix; Fibrillogenesis; Connective tissue Contents 1. Collagens—general introduction ............................................. 1532 2. Collagens—the basic structural module......................................... -

Sources of Collagen for Biomaterials in Skin Wound Healing
bioengineering Review Sources of Collagen for Biomaterials in Skin Wound Healing Evan Davison-Kotler 1,2, William S. Marshall 1 and Elena García-Gareta 2,* 1 Biology Department, St. Francis Xavier University, Antigonish, NS B2G 2W5, Canada 2 Regenerative Biomaterials Group, The RAFT Institute, Mount Vernon Hospital, Northwood HA6 2RN, UK * Correspondence: [email protected]; Tel.: +44-(0)-1923844350 Received: 23 May 2019; Accepted: 26 June 2019; Published: 30 June 2019 Abstract: Collagen is the most frequently used protein in the fields of biomaterials and regenerative medicine. Within the skin, collagen type I and III are the most abundant, while collagen type VII is associated with pathologies of the dermal–epidermal junction. The focus of this review is mainly collagens I and III, with a brief overview of collagen VII. Currently, the majority of collagen is extracted from animal sources; however, animal-derived collagen has a number of shortcomings, including immunogenicity, batch-to-batch variation, and pathogenic contamination. Recombinant collagen is a potential solution to the aforementioned issues, although production of correctly post-translationally modified recombinant human collagen has not yet been performed at industrial scale. This review provides an overview of current collagen sources, associated shortcomings, and potential resolutions. Recombinant expression systems are discussed, as well as the issues associated with each method of expression. Keywords: collagen; collagen sources; recombinant collagen; biomaterials; regenerative medicine; tissue engineering; wound healing; skin 1. Introduction 1.1. The Molecular Structure of Collagen The 28 different collagen types of the collagen superfamily are further divided into eight subfamilies, with the majority of the collagen types belonging to the fibril-forming or fibrillar subfamily [1,2]. -

Osteogenesis Imperfecta Types I-XI Implications for the Neonatal Nurse Jody Womack , RNC-NIC, NNP-BC, MS
Ksenia Zukowsky, PhD, APRN, NNP-BC ❍ Section Editor Beyond the Basics 3.0 HOURS Continuing Education Osteogenesis Imperfecta Types I-XI Implications for the Neonatal Nurse Jody Womack , RNC-NIC, NNP-BC, MS ABSTRACT Osteogenesis imperfecta (OI), also called “brittle bone disease,” is a rare heterozygous connective tissue disorder that is caused by mutations of genes that affect collagen. Osteogenesis imperfecta is characterized by decreased bone mass, bone fragility, and skin hyperlaxity. The phenotype present is determined according to the mutation on the affected gene as well as the type and location of the mutation. Osteogenesis imperfecta is neither preventable nor treatable. Osteogenesis imperfecta is classified into 11 types to date, on the basis of their clinical symptoms and genetic components. This article discusses the definition of the disease, the classifications on the basis of its clinical features, incidence, etiology, and pathogenesis. In addition, phenotype, natural history, diagnosis and manage- ment of this disease, recurrence risk, and, most importantly, the implications for the neonatal nurse and management for the family are discussed. Key Words: brittle bone disease , COL1A1 , COL1A2 , collagen disorders , osteogenesis imperfecta steogenesis imperfecta (OI) is a rare connec- is imperative to help the patient and the parents car- tive tissue disorder that is caused by muta- ing for infants born with OI. tions of genes that affect collagen. 1-4 Collagen O is the major protein of connective tissues, which is the REVIEW OF LITERATURE framework of bones. When collagen is not function- ing properly or there is lack of collagen in the tissue, Osteogenesis imperfecta was thought to have affected bones break easily.