Facets of Mechanical Regulation in the Morphogenesis of Craniofacial Structures
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
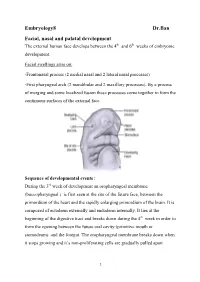
Embryology8 Dr.Ban Facial, Nasal and Palatal Development the External Human Face Develops Between the 4Th and 6Th Weeks of Embryonic Development
Embryology8 Dr.Ban Facial, nasal and palatal development The external human face develops between the 4th and 6th weeks of embryonic development. Facial swellings arise on: -Frontonasal process (2 medial nasal and 2 lateral nasal processes) -First pharyngeal arch (2 mandibular and 2 maxillary processes). By a process of merging and some localized fusion these processes come together to form the continuous surfaces of the external face. Sequence of developmental events : During the 3rd week of development an oropharyngeal membrane (buccopharyngeal ) is first seen at the site of the future face, between the primordium of the heart and the rapidly enlarging primordium of the brain. It is composed of ectoderm externally and endoderm internally. It lies at the beginning of the digestive tract and breaks down during the 4th week in order to form the opening between the future oral cavity (primitive mouth or stomodeum) and the foregut. The oropharyngeal membrane breaks down when it stops growing and it’s non-proliferating cells are gradually pulled apart 1 because they cannot fill the expanding area.The tissues around it expand very rapidly. The face develops from five primordia that appear in the fourth week: the frontonasal prominence, the two maxillary swellings, and the two mandibular swellings. The external face forms from two sources that surround the oropharyngeal membrane 1-Tissues of the frontonasal process that cover the forebrain, predominantly of neural crest origin. 2-The tissues of the first (or mandibular) pharyngeal arch, of mixed mesoderm and neural crest origin Face initially formed by 5 mesenchymal swellings ( prominences): Two mandibular prominences Two maxillary prominences Frontonasal prominence (midline structure is a single structure that is ventral to the forebrain. -

Anatomical Basis of Craniofacial Birth Defects
AnatomicalAnatomical BasisBasis ofof CraniofacialCraniofacial BirthBirth DefectsDefects Handout download: http://www.oucom.ohiou.edu/dbms-witmer/peds-rpac.htm LawrenceLawrence M.M. Witmer,Witmer, PhDPhD Department of Biomedical Sciences College of Osteopathic Medicine Ohio University Athens, Ohio 45701 [email protected] 18 October 2000 DevelopmentDevelopment ofof thethe FaceFace II • 5 facial primordia • Frontonasal prominence • Paired maxillary prominences • Paired mandibular prominences • Surround primordial mouth (stomodeum) • Neural crest: source for almost all connective tissues in the face • Frontonasal prominence forms forehead and nose and a short margin of mouth • Lower jaw and lip form first • Nasal placodes (and pit): surrounded by medial & lateral nasal prominences • Nasal pit remains connected to mouth • Maxillary prominences grow toward each other, pushing nasal prominences medially From Moore 1982 DevelopmentDevelopment ofof thethe FaceFace IIII • Medial nasal prominences merge with each other and with lateral nasal & maxillary prominences • Nasolacrimal groove: between lateral nasal and maxillary prominences • Becomes nasolacrimal duct • Duct forms as solid epithelial cord that later canalizes • Nasolacrimal duct atresia • Failure to completely canalize • 6% of newborns • Intermaxillary segment • Merger of medial nasal prominences • Gives rise to philtrum, premaxillary bones, primary palate From Moore 1982 SummarySummary ofof FacialFacial DevelopmentDevelopment From Moore 1982 Disruptions in the formation -

Development and Malformations of Face and Palate
Development and malformations of face and palate Compiled by András Csillag and Andrea D. Székely GERMINAL LAYER DERIVATIVES ECTODERM contributing to the formation of the face appears by the 4th week. The oropharyngeal membrane (interface between ECTODERM and ENDODERM) is located in front of the later palatine tonsils. Ectodermal structures limiting the stomodeum participate in the formation of the face, as well as of the nasal and oral cavities. MESENCHYME that fills the pharyngeal arches derives from the neural crest ECTOMESENCHYME DEVELOPMENT OF THE FACE WEEK 6 The face is formed by 5 processes approximately 24 days Frontonasal prominence (1) Maxillary prominence (2) 1st arch Mandibular prominence (2) nasal (olfactory) pits form surrounded by the medial and lateral nasal processes nasolacrimal groove separates the lateral nasal process from the maxillary process maxillary processes fuse with the medial nasal processes lateral nasal processes fuse with the maxillary processes, thus obliterating the nasolacrimal groove. DEVELOPMENT OF THE FACE 5-week embryo 6-week embryo 7-week embryo. 10-week embryo The nasal prominences are gradually Maxillary prominences have fused separated from the maxillary with the medial nasal prominences. prominence by deep furrows. CEPHALIC PRIMORDIA - PLACODES Olfactory placode Pharyngeal Stomodeum” arches Nasal placodes - thickenings of the surface ectoderm (later differentiate into the olfactory epithelium) DEVELOPMENT OF THE NASAL CAVITIES AND THE HARD PALATE By week 5 the placodes form the nasal pits. They further invaginate and the pits approach the primitive oral cavity. A thin oronasal membrane separates the two cavities. By its rupture the primitive choanae will form DEVELOPMENT OF THE NASAL CAVITIES AND THE HARD PALATE By week 8, a partition forms between the primitive nasal chambers and the oral cavity The primary palate - the anterior aspect derives from the intermaxillary segment (or median palatine process, formed by the medial nasal processes). -

A Combined Approach of Teaching Head Development Using Embryology and Comparative Anatomy
Edorium J Anat Embryo 2016;3:17–27. Danowitz et al. 17 www.edoriumjournals.com/ej/ae REVIEW ARTICLE PEER REVIEWED | OPEN ACCESS A combined approach of teaching head development using embryology and comparative anatomy Melinda Danowitz, Hong Zheng, Adriana Guigova, Nikos Solounias ABSTRACT the evolutionary changes of many structures allows for a greater understanding of the Many aspects of human head embryology reflect human embryology, and removes the need for its evolutionary development. The pharyngeal memorization of seemingly complex processes. arches, a major component of head development, A link to comparative evolutionary anatomy originally functioned in filter feeding and provides context to the purpose and morphology vascular exchange, which is why each arch of primitive structures, and clarifies several has associated vasculature and muscles. The issues in human head development. primitive tongue had few-associated muscles and was responsible for simple movements; the Keywords: Anatomy education, Embryology, Head human tongue evolved post-otic somites that and neck, Pharyngeal arches migrate to the tongue and develop the majority of the tongue musculature. These somites originate How to cite this article outside the tongue, and the motor innervation therefore differs from the general and special Danowitz M, Zheng H, Guigova A, Solounias N. A sensory innervation. In the primitive condition, combined approach of teaching head development the trapezius and sternocleidomastoid belonged using embryology and comparative anatomy. to a single muscle group that were involved in Edorium J Anat Embryo 2016;3:17–27. gill movements; they separate into two muscles with the reduction of certain skeletal elements, but retain the same innervation. -

Syndromes of the First and Second Branchial Arches, Part 1: Embryology and Characteristic REVIEW ARTICLE Defects
Syndromes of the First and Second Branchial Arches, Part 1: Embryology and Characteristic REVIEW ARTICLE Defects J.M. Johnson SUMMARY: A variety of congenital syndromes affecting the face occur due to defects involving the G. Moonis first and second BAs. Radiographic evaluation of craniofacial deformities is necessary to define aberrant anatomy, plan surgical procedures, and evaluate the effects of craniofacial growth and G.E. Green surgical reconstructions. High-resolution CT has proved vital in determining the nature and extent of R. Carmody these syndromes. The radiologic evaluation of syndromes of the first and second BAs should begin H.N. Burbank first by studying a series of isolated defects: CL with or without CP, micrognathia, and EAC atresia, which compose the major features of these syndromes and allow more specific diagnosis. After discussion of these defects and the associated embryology, we proceed to discuss the VCFS, PRS, ACS, TCS, Stickler syndrome, and HFM. ABBREVIATIONS: ACS ϭ auriculocondylar syndrome; BA ϭ branchial arch; CL ϭ cleft lip; CL/P ϭ cleft lip/palate; CP ϭ cleft palate; EAC ϭ external auditory canal; HFM ϭ hemifacial microsomia; MDCT ϭ multidetector CT; PRS ϭ Pierre Robin sequence; TCS ϭ Treacher Collins syndrome; VCFS ϭ velocardiofacial syndrome adiographic evaluation of craniofacial deformities is nec- major features of the syndromes of the first and second BAs. Ressary to define aberrant anatomy, plan surgical proce- Part 2 of this review discusses the syndromes and their radio- dures, and evaluate the effects of craniofacial growth and sur- graphic features: PRS, HFM, ACS, TCS, Stickler syndrome, gical reconstructions.1 The recent rapid proliferation of and VCFS. -

Illustrated Review of the Embryology and Development of the Facial
REVIEW ARTICLE Illustrated Review of the Embryology and Development of the Facial Region, Part 2: Late Development of the Fetal Face and Changes in the Face from the Newborn to Adulthood P.M. Som and T.P. Naidich ABSTRACT SUMMARY: The later embryogenesis of the fetal face and the alteration in the facial structure from birth to adulthood have been reviewed. Part 3 of the review will address the molecular mechanisms that are responsible for the changes described in parts 1 and 2. art 1 of this 3-part review primarily dealt with the early em- first make contact, each is completely covered by a homoge- Pbryologic development of the face and nasal cavity. Part 2 will neous epithelium. A special epithelium arises at the edge of discuss the later embryonic and fetal development of the face, and each palatal shelf, facilitating the eventual fusion of these changes in facial appearance from neonate to adulthood will be shelves. The epithelium on the nasal cavity surface of the palate reviewed. will differentiate into columnar ciliated epithelium. The epi- thelium on the oral cavity side of the palate will differentiate Formation of the Palate into stratified squamous epithelium. Between the sixth and 12th weeks, the palate is formed from 3 The 2 palatal shelves also fuse with the triangular primary pal- primordia: a midline median palatine process and paired lateral ate anteromedially to form a y-shaped fusion line. The point of palatine processes (Fig 1). In the beginning of the sixth week, fusion of the secondary palatal shelves with the primary palate is merging of the paired medial nasal processes forms the intermax- marked in the adult by the incisive foramen. -

EMBRYOLOGY7 Dr.Ban A.Ghani Developmdent of External
EMBRYOLOGY7 Dr.Ban A.Ghani EMBRYOLOGY7 Dr.Ban A.Ghani Developmdent of external /Sequence of developmental During the third week of development an oropharyngeal membrane buccopharyngeal , or oral membrane) is first seen at the site of the future face, between the primordium of the heart and the rapidly enlarging primordium of the brain. It is composed of ectoderm externally and endoderm internally. It lies at the beginning of the digestive tract and breaks down during the 4th week in order to form the opening between the future oral cavity (primitive mouth or stomodeum) and the foregut. The oropharyngeal membrane breaks down when it stops growing. While tissues around it expand very rapidly, the oropharyngeal membrane’s non-proliferating cells are gradually pulled apart because they cannot fill the expanding area. The human face begins to form during the 4th week of embryonic development. By the 6th week the external face is completed. The development of the palate subdivides nasal and oral cavities. This development continues into the 12th week with completion of the soft palate. Human face during the 4th prenatal week. Around the centrally located oral pit are grouped the frontal and maxillary processes and the mandibular arch. 1 EMBRYOLOGY7 Dr.Ban A.Ghani Human face during the 5th prenatal week. The nasal pits develop and appear on the sides of the face. The frontal process now becomes the frontonasal process Human face during the 6th prenatal week. Nasal pits appear more centrally located in the medial nasal process. This is the result of growth of the lateral face, which also causes the eyes to approach the front of the face. -

A New Origin for the Maxillary Jaw
Developmental Biology 276 (2004) 207–224 www.elsevier.com/locate/ydbio A new origin for the maxillary jaw Sang-Hwy Leea, Olivier Be´dardb,1, Marcela Buchtova´b, Katherine Fub, Joy M. Richmanb,* aDepartment of Oral, Maxillofacial Surgery and Oral Science Research Center, Medical Science and Engineering Research Center, BK 21 Project for Medical Science, College of Dentistry Yonsei University, Seoul, Korea bDepartment of Oral Health Sciences, Faculty of Dentistry, University of British Columbia, Vancouver, BC, Canada, V6T 1Z3 Received for publication 7 April 2004, revised 5 August 2004, accepted 31 August 2004 Available online 5 October 2004 Abstract One conserved feature of craniofacial development is that the first pharyngeal arch has two components, the maxillary and mandibular, which then form the upper and lower jaws, respectively. However, until now, there have been no tests of whether the maxillary cells originate entirely within the first pharyngeal arch or whether they originate in a separate condensation, cranial to the first arch. We therefore constructed a fate map of the pharyngeal arches and environs with a series of dye injections into stage 13–17 chicken embryos. We found that from the earliest stage examined, the major contribution to the maxillary bud is from post-optic mesenchyme with a relatively minor contribution from the maxillo-mandibular cleft. Cells labeled within the first pharyngeal arch contributed exclusively to the mandibular prominence. Gene expression data showed that there were different molecular codes for the cranial and caudal maxillary prominence. Two of the genes examined, Rarb (retinoic acid receptor b) and Bmp4 (bone morphogenetic protein) were expressed in the post-optic mesenchyme and epithelium prior to formation of the maxillary prominence and then were restricted to the cranial half of the maxillary prominence. -

Facial and Palatal Development
11. FACIAL AND PALATAL DEVELOPMENT Letty Moss-Salentijn DDS, PhD Dr. Edwin S.Robinson Professor of Dentistry (in Anatomy and Cell Biology) E-mail: [email protected] READING ASSIGNMENT: Larsen 3rd edition: p.352; pp.365-371; 398-404. SUMMARY: The external human face develops between the 4th and 6th weeks of embryonic development. Facial swellings arise on the frontonasal process (2 medial nasal and 2 lateral nasal processes) and the first pharyngeal arch (2 mandibular and 2 maxillary processes). By a process of merging and some localized fusion these processes come together to form the continuous surfaces of the external face. The primary palate is formed in this period by fusion/merging of the medial nasal and maxillary processes. Subsequently, between 6th and 12th embryonic/fetal weeks, the secondary palate is formed as the result of fusion between palatal processes growing from the oral surfaces of the maxillary processes. Each merging and fusion site is also the site of a potential facial or palatal cleft. LEARNING OBJECTIVES You should be able to: a. Demonstrate on a frontal image of a human face those parts of the face that are formed by contributions from the frontonasal process and those that are formed by contributions from the first pharyngeal arch. b. Describe the following as to site, composition, time of appearance, and fate: oropharyngeal membrane, oronasal membrane. c. List the derivatives of the four pairs of facial processes and the pair of palatal processes. d. Describe the development of the nose and primary palate. e. Explain the differences between the processes of merging and fusion. -

Orthodontic Management of Unilateral Cleft Lip and Palate KARIN
Orthodontic Management of Unilateral Cleft Lip and Palate KARIN VARGERVIK, D.D.S. San Francisco, California 94143 Experimental and clinical findings indicate that unfavorable adaptations of normal structures occur in children with clefts. It is postulated that these deviations from normal development are reversible and can be corrected or prevented by properly designed treatment. On this basis, five hypotheses were formulated and tested on the data from the subjects included in this study. The subjects consisted of 1) 16 children without clefts, 2) eight children who had complete unilateral clefts of the lip and palate but who had not received orthodontic treatment, and 3) 16 children who had complete unilateral clefts of the lip and palate and who had been treated by the described orthodontic procedures. The mean age for each group was 16 years. It was concluded that orthodontic treatment can be designed to: 1) counteract the forces which inhibit development of the maxillary alveolar process horizontally and vertically, 2) partially prevent the reduction in the forward growth of the maxilla, 3) provide adequate jaw and dental arch relationships, and 4) establish and maintain correct position of the maxillary segments. KEY WORDS: Cleft palate, cleft lip, orthodontics Introduction should be brought into positions that mini- mize the effect of their inadequacies. Craniofacial growth and development in A clear distinction between the primary individuals with clefts of the lip and palate and secondary effects of clefts on the facial are influenced by irregularities in embryonic skeleton and soft tissues cannot be made in development, which cause tissue deficiencies, children. An experimental model was, there- and by lip and palate surgery, which results fore, developed in our Center and has been in scar tissue and contractures. -

Pharyngeal Arches. Pharyngeal Pouches
Multimedial Unit of Dept. of Anatomy Jagiellonian University The head and neck regions of a 4-week human embryo somewhat resemble these regions of a fish embryo of a comparable stage of development. This explains the former use of the designation „branchial apparatus” – the adjective „branchial” is derived from the Greek word branchia – the gill. The pharyngeal apparatus consists of: pharyngeal arches pharyngeal pouches pharyngeal grooves pharyngeal membranes The pharyngeal arches begin to develop early in the fourth week as neural crest cells migrate into the future head and neck regions. Drawings illustrating the human pharyngeal apparatus. The first pharyngeal arch (mandibular arch) develops two prominences the maxillary prominence (gives rise to maxilla, zygomatic bone, and squamous part of temporal bone) the mandibular prominence (forms the mandible) Drawings illustrating the human pharyngeal apparatus. Drawings illustrating the human pharyngeal apparatus. Drawings illustrating the human pharyngeal apparatus. Drawing of the head, neck, and thoracic regions of a human embryo (about 28 days), illustrating the pharyngeal apparatus. During the fifth week, the second pharyngeal arch enlarges and overgrows the third and fourth arches, forming an ectodermal depression – the cervical sinus. A - Lateral view of the head, neck, and thoracic regions of an embryo (about 32 days), showing the pharyngeal arches and cervical sinus. B - Diagrammatic section through the embryo at the level shown in A, illustrating growth of the second arch over -

The Correlation of Chin Prominence and Esthetics
Graduate Theses, Dissertations, and Problem Reports 2015 The correlation of chin prominence and esthetics Nicholas Maddux Follow this and additional works at: https://researchrepository.wvu.edu/etd Recommended Citation Maddux, Nicholas, "The correlation of chin prominence and esthetics" (2015). Graduate Theses, Dissertations, and Problem Reports. 6137. https://researchrepository.wvu.edu/etd/6137 This Thesis is protected by copyright and/or related rights. It has been brought to you by the The Research Repository @ WVU with permission from the rights-holder(s). You are free to use this Thesis in any way that is permitted by the copyright and related rights legislation that applies to your use. For other uses you must obtain permission from the rights-holder(s) directly, unless additional rights are indicated by a Creative Commons license in the record and/ or on the work itself. This Thesis has been accepted for inclusion in WVU Graduate Theses, Dissertations, and Problem Reports collection by an authorized administrator of The Research Repository @ WVU. For more information, please contact [email protected]. CORRELATION OF CHIN PROMINENCE AND ESTHETICS Nicholas Maddux D.D.S. A THESIS Submitted to: The School of Dentistry At West Virginia University In partial fulfillment of the requirements For the degree of Master of Science In Orthodontics Tim Tremont D.M.D., M.S. Advisor Peter Ngan, D.M.D., Chair Chris Martin, D.D.S., M.S. Richard Jurevic, D.M.D, M.S. West Virginia University Department of Orthodontics Morgantown, West Virginia The Correlation of Chin Prominence and Facial Esthetics Nicholas Maddux, D.D.S., Dr.