An Introduction to the Biocomplexity of Sanak Island, Western Gulf of Alaska1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Darwin's Legacy
Darwin’s Darwin’s Legacy: Legacy: 24 TheThe Status Status of Evolutionary of Evolutionary ArchaeologyArchaeology in Argentina in Argentina Tribute to the 200th anniversary of the Editedbirth by of Charles Darwin and the 150th Marceloanniversary Cardillo & ofHernán the publication Muscio of The Origin of Species El Legado de Darwin: El estado de la Arqueología Evolucionista en Argentina Homenaje a los 200 años del nacimiento de Charles Darwin y a los 150 años de la publicación de El Origen de las Especies Access Archaeology o hae pre rc s A s A y c g c e o l s o s e A a r c Ah Edited by Marcelo Cardillo & Hernán Muscio Archaeopress Publishing Ltd Gordon House 276 Banbury Road Oxford OX2 7ED www.archaeopress.com ISBN 978 1 78491 270 3 (e-Pdf) ISBN 978 1 78491 276 5 © Archaeopress and the individual authors 2016 South American Archaeology Series No 24 Series Editor Andrés D. Izeta All rights reserved. No part of this book may be reproduced or transmitted, in any form or by any means, electronic, mechanical, photocopying or otherwise, without the prior written permission of the copyright owners. Contents PREFACE ������������������������������������������������������������������������������������������������������������������������������������� v Hernán Muscio, Marcelo Cardillo Introduction: Evolutionary Archaeology a comprehensive framework �������������������������������������� vii Hernán MUSCIO, Marcelo CARDILLO References �����������������������������������������������������������������������������������������������������������������������������������xiii -

500 Natural Sciences and Mathematics
500 500 Natural sciences and mathematics Natural sciences: sciences that deal with matter and energy, or with objects and processes observable in nature Class here interdisciplinary works on natural and applied sciences Class natural history in 508. Class scientific principles of a subject with the subject, plus notation 01 from Table 1, e.g., scientific principles of photography 770.1 For government policy on science, see 338.9; for applied sciences, see 600 See Manual at 231.7 vs. 213, 500, 576.8; also at 338.9 vs. 352.7, 500; also at 500 vs. 001 SUMMARY 500.2–.8 [Physical sciences, space sciences, groups of people] 501–509 Standard subdivisions and natural history 510 Mathematics 520 Astronomy and allied sciences 530 Physics 540 Chemistry and allied sciences 550 Earth sciences 560 Paleontology 570 Biology 580 Plants 590 Animals .2 Physical sciences For astronomy and allied sciences, see 520; for physics, see 530; for chemistry and allied sciences, see 540; for earth sciences, see 550 .5 Space sciences For astronomy, see 520; for earth sciences in other worlds, see 550. For space sciences aspects of a specific subject, see the subject, plus notation 091 from Table 1, e.g., chemical reactions in space 541.390919 See Manual at 520 vs. 500.5, 523.1, 530.1, 919.9 .8 Groups of people Add to base number 500.8 the numbers following —08 in notation 081–089 from Table 1, e.g., women in science 500.82 501 Philosophy and theory Class scientific method as a general research technique in 001.4; class scientific method applied in the natural sciences in 507.2 502 Miscellany 577 502 Dewey Decimal Classification 502 .8 Auxiliary techniques and procedures; apparatus, equipment, materials Including microscopy; microscopes; interdisciplinary works on microscopy Class stereology with compound microscopes, stereology with electron microscopes in 502; class interdisciplinary works on photomicrography in 778.3 For manufacture of microscopes, see 681. -

Paleozoology in the Service of Conservation Biology
Evolutionary Anthropology 15:11–19 (2006) ARTICLES Paleozoology in the Service of Conservation Biology R. LEE LYMAN Paleozoological data reveal past conditions created by anthropogenic and nat- grapple with the fact that ecosystems ural processes. These conditions can serve as benchmarks of ecological proper- and landscapes are not static for natu- ties and processes desired by conservation biologists. Paleozoological data pro- ral and anthropogenic reasons.16 Their vide empirical evidence analogous to experimental results of anthropogenic and desire to manage a minimally anthro- environmental causes. They can be used to determine whether a taxon is native or pogenically influenced ecosystem intro- exotic to an area, distinguish invasive from recolonizing taxa, choose a manage- duces the difficulty of identifying the ment action likely to produce a desired result, test benchmarks based on historic boundary between natural and unnatu- data, reveal unanticipated effects of conservation efforts, and identify causes of ral.17–19 But nonanthropogenically in- ecological conditions. It is time to use paleoecological knowledge in the service of fluenced ecosystems are not always modern conservation biology. desired. For example, some anthropo- genically introduced exotic taxa such as game birds in the western United States Conservation biologists, restoration compositions of biological communi- are economically beneficial and ecolog- ecologists, and wildlife managers of- ties occupying tens to hundreds of ically benign. ten select an ecological benchmark,1,2 hectares, as well as to ecosystems con- The paleozoological record pro- ecological baseline,3 or historical sisting of organisms, geology, fire re- vides unprecedented data that reflect landscape4,5 that they seek to recreate gimes, and so on, as well as ecological the long-term operation of many eco- or maintain in an area. -

AS General Studies to BS Biology, 4-8 Science Certification
Northeast Texas Community College & Texas A&M University – Texarkana 2019-2020 Guided Pathways Associate of Science in General Studies to Bachelor of Science in Biology, 4-8 Science Certification NTCC A&M-TEXARKANA COURSES HOURS COURSES HOURS ENGL 1301 3 BIOL 307 4 SPCH 1315 (or) SPCH 1321 3 BIOL 308 3 MATH 1314* 3 BIOL 310 4 BIOL 1406 4 BIOL 402 4 BIOL 1407 4 BIOL 466 4 HIST 2321* 3 BIOL 472 3 ARTS 1301* 3 UD Biology Electives 9 HIST 1301 3 RDG 343 3 HIST 1302 3 RDG 350 3 GOVT 2305 3 ED 311 3 GOVT 2306 3 ED 321 3 PSYC 2301* 3 BLOCK I ENGL 1302 3 ED 331 3 GEOL 1403 (CAO B) 4 ED 495 3 SPAN 1411* 4 BLOCK II PHED* 2 ED 496 3 MATH 1350 (Elective) 3 SPED 418 3 MATH 1351 (Elective) 3 PHYS 1415 (Elective) 4 PHYS 1417 (Elective) 4 2.8 Minimum GPA TOTAL 65 TOTAL 120 *Other Courses may Apply. See NTCC Degree Plan for Options 54 Upper Division (UD) Hours Required for the BS Degree 30 Hours of Residency Required Travel to Main Campus in Texarkana Required to Complete Degree UD= Upper Division LD= Lower Division Effective September 1, 2019 – August 31, 2024. This unofficial degree plan is for informational purposes only. Please contact [email protected] for questions. TEACHER PREPARATION PROGRAM ADMISSION REQUIREMENTS Apply 3rd Year, 1st Semester 1. Application to Teacher Prep Program via TK20 in September or February 2. GPA requirement of 2.8 cumulative 3. Completion of ED 311, ED 321 and SPED 410 with grade C or above 4. -

PALEOZOOLOGY : INTRODUCTION Dr Poonam Kumari Dept of Zoology (Bsc PART III Paper VI )
PALEOZOOLOGY : INTRODUCTION Dr Poonam kumari Dept Of Zoology (BSc PART III Paper VI ) It is the branch of paleontology, paleobiology, or zoology dealing with the recovery and identification of multicellular animal remains from geological (or even archeological) contexts, and the use of these fossils in the reconstruction of prehistoric environments and ancient ecosystems. Paleozoology is derived from greek work i.e palaeon means "old" and zoon " means animal. Vertebrate Paleozoology Vertebrate paleozoology refers to the use of morphological, temporal, and stratigraphic data to map vertebrate history in evolutionary theory. Vertebrates are classified as a subphylum of Chordata, a phylum used to classify species adhering to a rod-shaped, flexible body type called a notochord. Classes of vertebrates listed in chronological order from oldest to most recent include heterostracans, osteostracans, coelolepid agnathans, acanthodians, osteichthyan fishes, chondrichthyan fishes, amphibians, reptiles, mammals, and birds. All vertebrates are studied under standard evolutionary generalizations of behavior and life process, although there is controversy over whether population can be accurately estimated from limited fossil resources. Evolutionary origins of vertebrates as well as the phylum Chordata have not been scientifically determined. Many believe vertebrates diverged from a common ancestor of chordates and echinoderms. This belief is well supported by the prehistoric marine creature Amphioxus. Amphioxus does not possess bone, making it an invertebrate, but it has common features with vertebrates including a segmented body and a notochord. This could imply that Amphioxus is a transitional form between an early chordate, echinoderm or common ancestor, and vertebrates. Quantitative Paleozoology Quantitative paleozoology is a process of taking a census of fossil types rather than inventory. -

Towards a Historical Ecology of Intertidal Foraging in the Mafia Archipelago
Towards a Historical Ecology of Intertidal Foraging in the Mafia Archipelago: Archaeomalacology and Implications for Marine Resource Management Authors: Patrick Faulkner, Matthew Harris, Othman Haji, Alison Crowther, Mark C. Horton, et. al. Source: Journal of Ethnobiology, 39(2) : 182-203 Published By: Society of Ethnobiology URL: https://doi.org/10.2993/0278-0771-39.2.182 BioOne Complete (complete.BioOne.org) is a full-text database of 200 subscribed and open-access titles in the biological, ecological, and environmental sciences published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Complete website, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/terms-of-use. Usage of BioOne Complete content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Downloaded From: https://bioone.org/journals/Journal-of-Ethnobiology on 23 Jun 2019 Terms of Use: https://bioone.org/terms-of-use Access provided by Max Planck Digital Library Journal of Ethnobiology 2019 39(2): 182–203 Towards a Historical Ecology of Intertidal Foraging in the Mafia Archipelago: Archaeomalacology and Implications for Marine Resource Management Patrick Faulkner1,2*, Matthew Harris3, Othman Haji4, Alison Crowther2,3, Mark C. Horton5, and Nicole L. Boivin2 Abstract. Understanding the timing and nature of human influence on coastal and island ecosystems is becoming a central concern in archaeological research, particularly when investigated within a historical ecology framework. -

Course Syllabus BIOL 443/543 Paleozoology, Fall 2019
Texas A&M University-Texarkana - Course Syllabus BIOL 443/543 Paleozoology, Fall 2019 Instructor: Dr. Ben Neuman Office Location: SCIT 318G Phone: (903) 334 6654 Email: [email protected] Course Description: This course looks at the evolution of modern animals by bringing together recent advances in genetics with the fossil record. This course will provide an evolutionary perspective on the origins of important groups of animals from single-celled organisms to modern humans through lectures, discussions and hands-on workshops with fossils. Prerequisites: Invertebrate zoology or permission from instructor. Course Delivery Method: Face to Face Textbooks/Resources: This course will also draw on papers available through the Texas A&M- Texarkana Library, which will be made available to students through Blackboard. You will also need some blank paper and pencils for lab work. If you are able, please bring a laptop or device capable of searching the internet and recording notes to class. If you do, please remember to bring a power supply. Useful books to pick up used if you want to learn more, but not required: Introduction to Paleobiology and the Fossil Record. Benton MJ, paperback ISBN: 978-1- 4051-4157-4 The Rise of Animals: Evolution and Diversification of the Kingdom Animalia. Mikhail A. Fedonkin, James G. Gehling, Kathleen Grey, Guy M. Narbonne, and Patricia Vickers-Rich Hardback, 344 pages. ISBN:9780801886799 January 2008 Student Learning Outcomes: 1. Learn to identify important groups of animals in the fossil record by synthesizing information about comparative anatomy and taphonomy 2. Analyze trace fossil data to better understand the behavior and ecology of extinct animals 3. -

Life Sciences Glossary: Fields of Biology: Malacology and Mammalogy- Examrace
9/17/2021 Life Sciences Glossary: Fields of Biology: Malacology and Mammalogy- Examrace Examrace Life Sciences Glossary: Fields of Biology: Malacology and Mammalogy Glide to success with Doorsteptutor material for competitive exams : get questions, notes, tests, video lectures and more- for all subjects of your exam. Fields of Biology Malacology: Study of mollusks. Mammalogy: Study of mammals. Mastology: Study of breast including teats. Melanology: Study of pigments. Molecular Biology: Study of life sciences on molecular level (i.e.. RNA and DNA level) . Mycology: Study of fungi. Myology (Sarcology) : Study of muscles. Myrmecology: Study of ants. Neonatology: Study of the new borns up to two months of age. Nephrology: Study of kidney. Neurology: Study of nervous system. Nidology: Study of nests of birds. Nosology: Study of diseases. Odontology: Study of teeth and gums. Olericulture: The study of vegetable yielding plants. Oncology: Study of cancer. Oneirology: Study of dreams. Ontogeny: Embryonic history. Oology: Study of eggs of birds. Ophthalmology: Study of eyes. Organocology: Study of development of organs under embryology. Organology: Study of organs. Ornithology: Study of birds. 1 of 4 9/17/2021 Life Sciences Glossary: Fields of Biology: Malacology and Mammalogy- Examrace Osteology: Study of bones. Oto-rhino-laryngology: Study of ear, nose and larynx. Pedology: Study of larval stages. Paleontology (Gr. Apples-ancient being) : The branch of science which deals with the study of fossils and their distribution in time. Paleozoology: The branch of science which deals with the study of fossils of animals. Palynology: Study of pollen grains in relation to taxonomy and evolution etc. Parasitology: Study of parasites. -
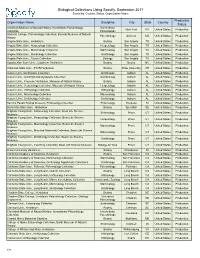
Biological Collections Using Specify, September 2017 Sorted By: Country, Status, Organization Name
Biological Collections Using Specify, September 2017 Sorted by: Country, Status, Organization Name Production Organization Name Discipline City State Country Status American Museum of Natural History, Invertebrate Paleontology Invertebrate New York NY United States Production Collection Paleontology Amherst College, Paleontology Collection, Beneski Museum of Natural Paleontology Amherst MA United States Production History Angelo State Univ., Herbarium Botany San Angelo TX United States Production Angelo State Univ., Herpetology Collection Herpetology San Angelo TX United States Production Angelo State Univ., Mammalogy Collection Mammalogy San Angelo TX United States Production Angelo State Univ., Ornithology Collection Ornithology San Angelo TX United States Production Angelo State Univ., Tissue Collection Zoology San Angelo TX United States Production Appalachian State Univ., Carpenter Herbarium Botany Boone NC United States Production Arkansas State Univ., STAR Herbarium Botany State University AR United States Production Auburn Univ. Ornithology Collection Ornithology Auburn AL United States Production Auburn Univ., Arachnid and Myriapods Collection Arachnology Auburn AL United States Production Auburn Univ., Freeman Herbarium, Museum of Natural History Botany Auburn AL United States Production Auburn Univ., Herpetology Collection, Museum of Natural History Herpetology Auburn AL United States Production Auburn Univ., Ichthyology Collection Ichthyology Auburn AL United States Production Auburn Univ., Mammalogy Collection Mammalogy Auburn -

Alfred Russel Wallace
ALFRED RUSSEL WALLACE By MARTIN FICHMAN1 /?y-/» Glendon College, York University, Toronto TWAYNE PUBLISHERS A DIVISION OF G. K. HALL & CO., BOSTON 1981 Contents About the Author Preface Chronology 1. Introduction 15 2. Natural Selection 29 3. Biogeography 60 4. Human Evolution 99 5. Social and Political Concerns 122 6. Conclusion 159 Notes and References 163 Selected Bibliography 174 Index 183 Alfred Russel Wallace About the Author Bom in Brooklyn, New York, in 1944, Martin Fichman received the B.Sc. degree from the Polytechnic Institute of New York and the A.M. and Ph.D. degrees from Harvard University and is currently associate professor of History and Natural Science at York University (Glendon College), Toronto. He has pub¬ lished articles on Alfred Russel Wallace and on the history of chemistry and is a contributor to the Dictionary of Scientific Biography. He is presently working on a study of the inter¬ relation between biological and sociopolitical ideas in the writings of late nineteenth-century evolutionists. Preface Alfred Russel Wallace was among the most brilliant Victorian naturalists and codiscoverer of one of the principal scientific achievements of the nineteenth century, the theory of natural selection. Although his accomplishments were fully recognized by his contemporaries, his reputation diminished somewhat in this century. Paradoxically, this situation has arisen partly through Wallace’s persistent efforts to equate evolution by natural selection with the name of Charles Darwin in the public mind. I have, therefore, critically analyzed Wallace’s major theoretical advances in order to clarify his central role in the history of evolutionary biology. Given the multiplicity of his scientific inter¬ ests, I have focused on one aspect—his biogeographical system— which best exemplifies the broad power of his evolutionary syn¬ thesis. -

Effect of Different Acidifying Agents on Amendment in Buffering Capacity Of
e-ISSN:2320-0189 Research & Reviews: Journal of Botanical Sciences p-ISSN:2347-2308 Brief Note on Ancient Paleobotany Han Asard* Department of Biology Molecular, Plant Physiology and Biotechnology, University of Antwerp, Belgium Editorial Note Received: 19/09/2020 *For Correspondence Accepted: 23/09/2020 Published: 30/09/2020 Han Asard, Department of Biology Molecular, Plant Physiology and Biotechnology, University of Antwerp, Belgium. E-mail: [email protected] Paleobotany is the investigation of the plants that lived quite a while in the past. The prefix paleo originates from the Greek word paleon which implies old, while, botany is the investigation of plants. It is one portion of a bigger part of science called Fossil Science (paleontology) which considers the historical backdrop of life on Earth more by and large. Paleobotany explicitly centers on the investigation of vegetation, while paleozoology centers on creature i.e. animal life. Paleobotany helps in responding to different inquiries, for example, what sorts of plants may a ravenous dinosaur have eaten? When did blossoms initially show up on Earth? How did plants add to the development of earth's present climate? What’s more, some more. Paleobotany permits us to think about the kinds of plants that lived sometime in the past. It encourages us to find out about what life on Earth resembled in the far off past. Paleobotany has a long history in the kingdom of science. Plant fossils are generally effectively unmistakable and individuals all through the world have been finding and gathering them for many years. By the mid-1700’s, a few books had just been distributed that included representations of plant fossils. -

AMERICAN MUSEUM of NATURAL HISTORY OFFICE of GRANTS and FELLOWSHIPS Central Park West at 79Th Street New York, New York 10024-5192
Last Name ________________________________ PLEASE PRINT AMERICAN MUSEUM OF NATURAL HISTORY OFFICE OF GRANTS AND FELLOWSHIPS Central Park West at 79th Street New York, New York 10024-5192 RESEARCH GRANTS APPLICATION for (please check one): q THEODORE ROOSEVELT MEMORIAL FUND Application deadline: February 15, Announcement: April 30 q LERNER-GRAY FUND FOR MARINE RESEARCH Application deadline: March 15, Announcement: May 15 INSTRUCTIONS A completed signed form and all supporting material (except letters of recommendation) should be submitted IN DUPLICATE to the attention of the appropriate Fund, American Museum of Natural History, Central Park West at 79th Street, New York, New York, 10024-5192. BRIEF DESCRIPTION OF FUNDS The Lerner-Gray Fund for Marine Research is intended for projects in marine zoology with emphasis on systematics, evolution, ecology, and field-oriented behavior; it does not support marine botany. The Theodore Roosevelt Memorial Fund is directed toward research on North American fauna, except birds. These funds support research in either neontology or paleontology. Awards are for one year and average $1,750; most grants range between $200-$2,500. Applicants are usually allowed a maximum of two awards. APPLICATION GUIDELINES · The budget on 1 separate page must show clearly the amounts and purposes for which the award will be used. If travel by automobile is required, an allowance of up to 40.5 cents per mile may be used in calculating costs. The following items are most commonly supported: Consumable supplies or expendable equipment, living expenses in the field or at a research station, and travel expenses. The following items are not supported: Permanent equipment, salary for principal investigator, overhead, conference or meeting costs, expenses and travel for field assistant.