Title of Thesis: ABSTRACT PHYSIOLOGICAL and BEHAVIORAL EFFECTS of HANDLING and RESTRAINT on the BALL PYTHON (Python Regius)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Quick Reference Guide: Introduced Constrictors in Florida1 Steve A
WEC302 Quick Reference Guide: Introduced Constrictors in Florida1 Steve A. Johnson and Monica E. McGarrity2 Three non-native species of large constrictor snakes are that these were escaped or released pets. View maps of loca- now breeding in Florida, and several others have been tions where each species has been encountered in Florida encountered but have not yet established wild populations. by visiting the EDDMapS Florida invasive species reporting This fact sheet, best viewed as a pdf (http://edis.ifas.ufl.edu/ portal online at http://www.IveGot1.org. Learn more about pdffiles/UW/UW34700.pdf), is a quick reference guide how to scan for, recognize, and report introduced constric- to identification of the constrictors you are most likely to tors by completing the Introduced Reptile Early Detection encounter in Florida. Although many of these snakes are and Documentation training course. Visit http://ufwildlife. not established in the wild, they are common in the pet ifas.ufl.edu/reddy.shtml to learn more and get REDDy! trade, and each has been spotted in the wild—it is likely Pythons Burmese Python (Python bivittatus) Status: established, breeding populations; range expanding Head: dark arrowhead, light center line, dark and light in Florida wedges under eyes Size: up to 12 feet or longer Body: Giraffe-like spots, dark blotches not connected Figure 1. Burmese python. Credits: Head illustration by USGS; body illustration by Monica E. McGarrity, UF 1. This document is WEC302, one of a series of the Department of Wildlife Ecology and Conservation, UF/IFAS Extension. Original publication date November 2010. Revised February 2014 and June 2017. -

A New Species of Hepatozoon (Apicomplexa: Adeleorina) from Python Regius (Serpentes: Pythonidae) and Its Experimental Transmission by a Mosquito Vector
J. Parasitol., 93(?), 2007, pp. 1189–1198 ᭧ American Society of Parasitologists 2007 A NEW SPECIES OF HEPATOZOON (APICOMPLEXA: ADELEORINA) FROM PYTHON REGIUS (SERPENTES: PYTHONIDAE) AND ITS EXPERIMENTAL TRANSMISSION BY A MOSQUITO VECTOR Michal Sloboda, Martin Kamler, Jana Bulantova´*, Jan Voty´pka*†, and David Modry´† Department of Parasitology, University of Veterinary and Pharmaceutical Sciences, Palacke´ho 1-3, 612 42 Brno, Czech Republic. e-mail: [email protected] ABSTRACT: Hepatozoon ayorgbor n. sp. is described from specimens of Python regius imported from Ghana. Gametocytes were found in the peripheral blood of 43 of 55 snakes examined. Localization of gametocytes was mainly inside the erythrocytes; free gametocytes were found in 15 (34.9%) positive specimens. Infections of laboratory-reared Culex quinquefasciatus feeding on infected snakes, as well as experimental infection of juvenile Python regius by ingestion of infected mosquitoes, were performed to complete the life cycle. Similarly, transmission to different snake species (Boa constrictor and Lamprophis fuliginosus) and lizards (Lepidodactylus lugubris) was performed to assess the host specificity. Isolates were compared with Hepatozoon species from sub-Saharan reptiles and described as a new species based on the morphology, phylogenetic analysis, and a complete life cycle. Hemogregarines are the most common intracellular hemo- 3 genera (Telford et al., 2004). Low host specificity of Hepa- parasites found in reptiles. The Hemogregarinidae, Karyolysi- tozoon spp. is supported by experimental transmissions between dae, and Hepatozoidae are distinguished based on the different snakes from different families. Ball (1967) observed experi- developmental patterns in definitive (invertebrate) hosts oper- mental parasitemia with Hepatozoon rarefaciens in the Boa ating as vectors; all 3 families have heteroxenous life cycles constrictor (Boidae); the vector was Culex tarsalis, which had (Telford, 1984). -

Dispelling Python Regius Myths "Gorzula, Stefan, William Owusu Nsiah, and William Oduro
Dispelling Python regius Myths "Gorzula, Stefan, William Owusu Nsiah, and William Oduro. Survey of the status and By Francis Cosquieri management of the Royal Python (Python regius) in Ghana. Secrétariat CITES, 1997." This paper lists several pythons being found in Before we get deep into the “nitty gritty” of the trees, although points out that the species is subject I want to take a moment to say that the very adaptable to the point of being AHH Royal Python sub-group "Not Just a Pet semi-invasive and responds well to Rock" is the best place to start for getting rid of anthropogenic disturbance. the preconceptions surrounding the poor old Royal Python. Zack Tippie and the other admins have done an incredible job of "Luiselli, Luca, and Francesco Maria Angelici. dispelling the myths surrounding Royal Python "Sexual size dimorphism and natural history behaviour and husbandry, and Zack’s care traits are correlated with intersexual dietary guide for this oft-maligned snake (to be found divergence in royal pythons (Python regius) in the third Advancing Herpetological from the rainforests of southeastern Nigeria." Husbandry Quarterly Newsletter) is one of the Italian Journal of Zoology 65.2 (1998): best I have read. 183-185." - In the meantime, here is a post I made some Half of the male pythons encountered over a time ago on AHH itself bringing together some two year period were found on trees, there is a information on the wild habits of this python. large discrepancy between habitat use by All the papers mentioned here can be found in male and female pythons, and by larger and the AHH Files section. -

Investigations Into the Presence of Nidoviruses in Pythons Silvia Blahak1, Maria Jenckel2,3, Dirk Höper2, Martin Beer2, Bernd Hoffmann2 and Kore Schlottau2*
Blahak et al. Virology Journal (2020) 17:6 https://doi.org/10.1186/s12985-020-1279-5 RESEARCH Open Access Investigations into the presence of nidoviruses in pythons Silvia Blahak1, Maria Jenckel2,3, Dirk Höper2, Martin Beer2, Bernd Hoffmann2 and Kore Schlottau2* Abstract Background: Pneumonia and stomatitis represent severe and often fatal diseases in different captive snakes. Apart from bacterial infections, paramyxo-, adeno-, reo- and arenaviruses cause these diseases. In 2014, new viruses emerged as the cause of pneumonia in pythons. In a few publications, nidoviruses have been reported in association with pneumonia in ball pythons and a tiger python. The viruses were found using new sequencing methods from the organ tissue of dead animals. Methods: Severe pneumonia and stomatitis resulted in a high mortality rate in a captive breeding collection of green tree pythons. Unbiased deep sequencing lead to the detection of nidoviral sequences. A developed RT-qPCR was used to confirm the metagenome results and to determine the importance of this virus. A total of 1554 different boid snakes, including animals suffering from respiratory diseases as well as healthy controls, were screened for nidoviruses. Furthermore, in addition to two full-length sequences, partial sequences were generated from different snake species. Results: The assembled full-length snake nidovirus genomes share only an overall genome sequence identity of less than 66.9% to other published snake nidoviruses and new partial sequences vary between 99.89 and 79.4%. Highest viral loads were detected in lung samples. The snake nidovirus was not only present in diseased animals, but also in snakes showing no typical clinical signs. -

Ball Python Care Sheet
Ball Python Care Sheet NATURAL HISTORY: The Ball Python (also called the Royal Python) is by far the most common pet python in this part of the world. Ball pythons are native to sub-Saharan West Africa and thrive in extreme heat and humidity. Ball pythons are generally hardy and tend to do well in captivity if all of their basic husbandry needs are met. They are friendly, easy to handle, and a bit shy — making them a favorite amongst many reptile owners. Males range in size from 2-3 feet and females 3-5 feet. A ball python’s lifespan is variable, however, they can live for up to 25-35 years in captivity. DIET: Ball pythons consume rodents exclusively. Ball pythons can be fed mice or rats depending on the age and size of the snake. As a rule of thumb, the size (largest diameter) of the prey item should be approximately the same diameter as the snake’s body. Feeder rodents can be purchased alive or frozen. I recommend feeding frozen-thawed or pre-killed rodents to all pet snakes because live prey items are able to fight back. Keep in mind that rodents have very strong jaws, with sharp teeth, and are capable of inflicting serious wounds on your pet. Therefore, a live rodent should never be left in a snake’s enclosure without direct supervision. Some people prefer to feed snakes in a separate bin or tank (outside of their primary enclosure) which may help prevent accidental bites while reaching into their primary enclosure. Juvenile ball pythons should be fed weekly and adults may be fed bi-weekly or monthly depending on the size of the prey item. -
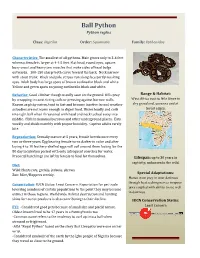
Ball Python Python Regius
Ball Python Python regius Class: Reptilia Order: Squamata Family: Pythonidae Characteristics: The smallest of all pythons. Male grows only to 3-4 feet whereas female is larger at 4-4.5 feet. Flat head, round eyes, square, boxy snout and heavy jaw muscles that make sides of head bulge outwards. 100-150 sharp teeth curve toward the back. Neck narrow with stout trunk. Black and pale stripes run along face partly masking eyes. Adult body has large spots of brown outlined in black and white. Yellow and green spots on young outlined in black and white. Behavior: Good climber though usually seen on the ground. Kills prey Range & Habitat: by wrapping in constricting coils or pressing against burrow walls. West Africa east to Nile River in Known as picky eaters, tend to fast and become inactive in cool weather dry grassland, savanna and at as bodies are not warm enough to digest food. Hisses loudly and coils forest edges. into tight ball when threatened with head and neck tucked away into middle. Hide in mammal burrows and other underground places. Eats weekly and sheds monthly with proper humidity. Captive adults rarely bite. Reproduction: Sexually mature at 5 years, female breeds once every two or three years. Egg bearing female turns darker in color and after laying 4 to 10 leathery-shelled eggs will coil around them fasting for the 80 day incubation period with only infrequent searches for water. Precocial hatchlings are left by female to fend for themselves. Lifespan: up to 30 years in captivity, unknown in the wild. -

THE HUSBANDRY and CAPTIVE PROPAGATION of the SOUTHERN ROCK PYTHON, PYTHON SEBAE NATALENSIS GERALD V HAAGNER Port Elizabeth Snake Park, P.O
BIMCI) I lerpciolopt.)1 Sotici.1 Both:NIL No. 42. 1992/93. THE HUSBANDRY AND CAPTIVE PROPAGATION OF THE SOUTHERN ROCK PYTHON, PYTHON SEBAE NATALENSIS GERALD V HAAGNER Port Elizabeth Snake Park, P.O. Box 13147 Hum wood, 6013. South Africa INTRODUCTION The African Rock Python, Python sebae (Fig. I) is distributed throughout much of sub-Saharan Africa, extending along the Nile valley into Egypt. In southern Africa it is restricted to northern Namibia and Botswana, extending through the northern and eastern Tansvaal, to Natal and the Transkei (Map 1). Pythons were previously found in the eastern Cape Province as far south as Bathurst where it became extinct in 1927 (Broadley, 1983). Branch (1988a) reported on an apparent successful reintroduction of the species into the eastern Cape. Broadley (1984) recently recognised the southern race (Python sebac natalensis) of the African Rock Python as distinct from the typical race from west and central Africa. The recently described Lesser Rock Python (P. saxuloides) from Kenya (Miller & Smith 1979), is treated as a junior synonym of the southern race (Broadley 1983). The African Rock Python is a thick-bodied snake with distinctive markings and size. As with most "giant" snakes. the size is easily over-exaggerated and most recorded lengths have been of the nominate race. Python sebae sebae has been reported to attain a length of 9.8m in the Ivory Coast (Doucet 1963), but full details for this specimen are lacking and cannot now be verified. Arthur Loveridge, whilst collecting in East Africa in 1927, measured a fresh skin of 9.1 metres and even allowing for extensive stretching, it must have been over seven metres long. -

G Iant Snakes
Copyrighted Material Some pages are omitted from this book preview. Giant Snakes Giant Giant Snakes A Natural History John C. Murphy & Tom Crutchfield Snakes, particularly venomous snakes and exceptionally large constricting snakes, have haunted the human brain for a millennium. They appear to be responsible for our excellent vision, as well as the John C. Murphy & Tom Crutchfield & Tom C. Murphy John anxiety we feel. Despite the dangers we faced in prehistory, snakes now hold clues to solving some of humankind’s most debilitating diseases. Pythons and boas are capable of eating prey that is equal to more than their body weight, and their adaptations for this are providing insight into diabetes. Fascination with snakes has also drawn many to keep them as pets, including the largest species. Their popularity in the pet trade has led to these large constrictors inhabiting southern Florida. This book explores what we know about the largest snakes, how they are kept in captivity, and how they have managed to traverse ocean barriers with our help. Copyrighted Material Some pages are omitted from this book preview. Copyrighted Material Some pages are omitted from this book preview. Giant Snakes A Natural History John C. Murphy & Tom Crutchfield Copyrighted Material Some pages are omitted from this book preview. Giant Snakes Copyright © 2019 by John C. Murphy & Tom Cructhfield All rights reserved. No part of this book may be reproduced in any form or by any electronic or mechanical means including information storage and retrieval systems, without permission in writing from the publisher. Printed in the United States of America First Printing March 2019 ISBN 978-1-64516-232-2 Paperback ISBN 978-1-64516-233-9 Hardcover Published by: Book Services www.BookServices.us ii Copyrighted Material Some pages are omitted from this book preview. -

Ball Pythons Purchased at Pet Stores and from Good Breeders in the U.S
Ball (Royal) Python Python regius LIFE SPAN: 20-30 years AVERAGE SIZE: 3-4 feet. Have grown as large as 5 feet * Females will grow larger than males CAGE TEMPS: Warm side – 85-90 0 F CAGE HUMIDITY: 50% Basking – 90 0 F * May need moist box for shedding time Cool side – 75 0 F * If room temperature falls below 75 at night, a supplemental infrared or ceramic heat fixture may be necessary. WILD HISTORY: Most ball pythons purchased at pet stores and from good breeders in the U.S. are bred and hatched in captivity. To help reduce the number of snakes poached from the wild; do make sure your pet is indeed captive bred. The “ball python” name comes from the manner in which they curl up into a ball when they are nervous. They will carefully place their heads in the middle of this tight ball for protection. The ball python species is originally from the forests of Central and Western Africa, and they enjoy both the ground and trees (semi-arboreal). In Europe they are known as “Royal Pythons” PHYSICAL CHARACTERISTICS: Ball pythons have flat heads and a square snout. They have 100-150 teeth that curve inward, which allow them to hold onto their prey. Normal color ball pythons have black striping on their heads and irregular brown spots on their bodies called “lobes”. “Pits” on the fronts of their face, formed by special scales, allow the snake to sense changes in heat as finite as 3/1000th of a degree. Ball pythons can also see extremely well in darkness. -

How to Keep Your Ball Python Healthy, Happy and Safe!
COMPANION REPTILE CARE SERIES Always Most Common Disorders of Ball Pythons llBuy from a reputable breeder because a young, captive-bred ball Intestinal parasites python is less stressed and more willing to eat than a frightened, llSkin problems wild-caught snake. llRespiratory disease llTake your new ball python to a reptile veterinarian for a general llMouth rot health exam and to test for parasites. llExternal parasites (mites and ticks) llLeave a newly purchased snake alone in its enclosure for 1-2 weeks llDysecdysis (retained skin shed, retained eye spectacles) to acclimate to its new home. llObstipation/constipation llProvide heat to your snake by special reptile heating pads or llBite wounds from prey incandescent light bulbs in reflector hoods; the use of “hot rocks” in llDystocia/retained eggs HOW TO the snake’s enclosure should be avoided. llRectal/colon or uterine prolapse llInteract with your ball python during the evening hours when llTrauma/thermal burns it is active. KEEP YOUR llUse a pillowcase or locking Tupperware container as a transport Ball pythons can flourish as long-lived pets when attention is container for short trips. given to certain aspects of husbandry, including temperature and humidity. Regular visits to your reptile veterinarian should be scheduled BALL PYTHON to check for parasites and other early signs of disease and to promote Housing for your Ball Python should: a long, satisfying relationship with your ball python. For help in finding llbe a relatively large enclosure. a reptile veterinarian in your area, contact the Association of Reptilian llmaintain ambient daytime temperatures of 80-85°F (27-29°C). -

Haemogregarine Genetic Diversity in Captive African Rock Pythons From
28 Original Investigation / Özgün Araştırma Haemogregarine Genetic Diversity in Captive African Rock Pythons from Nigeria Suggests a Geographical Pattern Nijerya’daki Afrika Kaya Pitonlarındaki Hemogregarin Genetik Çeşitlilik Coğrafik Bir Paterni Göstermektedir 1 2,3 2 4 Henry Olanrewaju Jegede , Beatriz Tomé , Ana Perera , Temidayo O. Omobowale 1Veterinary Teaching Hospital, University of Ilorin, Ilorin, Nigeria 2CIBIO, InBIO - Research Network in Biodiversity and Evolutionary Biology, Universidade do Porto, Campus de Vairão, Rua Padre Armando Quintas, Vairão, Portugal 3Departamento de Biologia, Faculdade de Ciências da Universidade do Porto, Rua do Campo Alegre, Porto, Portugal 4Department of Veterinary Medicine, University of Ibadan, Ibadan, Nigeria Cite this article as: Jegede HO, Tomé B, Perera A, Omobowale TO. Haemogregarine Genetic Diversity in Captive African Rock Pythons from Nigeria Suggests a Geographical Pattern. Turkiye Parazitol Derg 2018; 42:28-32. ABSTRACT Objective: Understanding the processes that drive parasite diversification, distribution, and abundance is central to disentangle the dynami- cs and evolution of diseases. In this study, we screened African rock pythons from Nigeria for the presence of blood parasites to assess their distribution, diversity, and phylogenetic relationships. Methods: A total of 21 captive African rock pythons collected from across 11 locations in Nigeria were sampled between August 2016 and January 2017. Samples were microscopically and genetically analyzed. Results: From the blood smears analyzed, 10 (47.6%) snakes were found to be infected with haemogregarines. Eight of the infected samples were genetically assessed and confirmed as haemogregarines of the recently describedBartazoon group. Two haplotypes were retrieved, of which one was distributed in the northern-central sampled localities and the other in the southern localities. -

The Role of Temperature and Humidity in Python Nest Site Selection
Animal Behaviour 81 (2011) 1077e1081 Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav The role of temperature and humidity in python nest site selection Z. R. Stahlschmidt*, J. Brashears, D. F. DeNardo School of Life Sciences, Arizona State University, Tempe article info Parental care is a convergent trait shown by a broad range of taxa. Often, successful parents must balance Article history: multiple developmental variables (e.g. embryonic water balance and thermoregulation). Pythons have Received 21 October 2010 recently emerged as valuable parental care models because females show simple egg-brooding behav- Initial acceptance 6 January 2011 iours that significantly influence variables of widespread importance (i.e. embryonic predation, hydra- Final acceptance 16 February 2011 tion, temperature and respiration). Nest site selection is an important parental behaviour that has been Available online 24 March 2011 shown to enhance several developmental variables in numerous taxa. In pythons, where brooding can MS. number: A10-00726 substantially mitigate environmental conditions to enhance the developmental environment, it is unclear to what extent females utilize environmental cues in selecting their nest site. Thus, we deter- Keywords: mined whether nest humidity and temperature influence python nest site selection because these fi adaptive signi cance variables influence python egg-brooding behaviour and are strongly associated with offspring fitness. We Antaresia childreni ’ created a radial maze with three nest site options: OTH: optimal temperature (31.5 C) and humidity Children s python 3 oviposition site selection (23 g/m H2O), as determined by previous studies; OT: optimal temperature, suboptimal humidity (13 g/ 3 parental care m H2O); OH: suboptimal temperature (25 C) and optimal humidity.