Biological Nomenclature, Classification and the Ethnozoological Specieme
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Linn E and Taxonomy in Japan: on the 300Th Anniversary of His Birth
No. 3] Proc. Jpn. Acad., Ser. B 86 (2010) 143 Linne and taxonomy in Japan: On the 300th anniversary of his birth By Akihito (His Majesty The Emperor of Japan) (Communicated by Koichiro TSUNEWAKI, M.J.A.) President, dear friends bers of stamens belonged to dierent classes, even when their other characteristics were very similar, I am very grateful to the Linnean Society of while species with the same number of stamens be- London for the kind invitation it extended to me to longed to the same class, even when their other participate in the celebration of the 300th anniver- characteristics were very dierent. This led to the sary of the birth of Carl von Linne. When, in 1980, I idea that the classication of organisms should be was elected as a foreign member of the Society, I felt based on a more comprehensive evaluation of all I did not really deserve the honour, but it has given their characteristics. This idea gained increasing me great encouragement as I have tried to continue support, and Linne’s classication system was even- my research, nding time between my ofcial duties. tually replaced by systems based on phylogeny. Today, I would like to speak in memory of Carl The binomial nomenclature proposed by Linne, von Linne, and address the question of how Euro- however, became the basis of the scientic names of pean scholarship has developed in Japan, touching animals and plants, which are commonly used in the upon the work of people like Carl Peter Thunberg, world today, not only by people in academia but also Linne’s disciple who stayed in Japan for a year as by the general public. -

Botanical Nomenclature: Concept, History of Botanical Nomenclature
Module – 15; Content writer: AvishekBhattacharjee Module 15: Botanical Nomenclature: Concept, history of botanical nomenclature (local and scientific) and its advantages, formation of code. Content writer: Dr.AvishekBhattacharjee, Central National Herbarium, Botanical Survey of India, P.O. – B. Garden, Howrah – 711 103. Module – 15; Content writer: AvishekBhattacharjee Botanical Nomenclature:Concept – A name is a handle by which a mental image is passed. Names are just labels we use to ensure we are understood when we communicate. Nomenclature is a mechanism for unambiguous communication about the elements of taxonomy. Botanical Nomenclature, i.e. naming of plants is that part of plant systematics dealing with application of scientific names to plants according to some set rules. It is related to, but distinct from taxonomy. A botanical name is a unique identifier to which information of a taxon can be attached, thus enabling the movement of data across languages, scientific disciplines, and electronic retrieval systems. A plant’s name permits ready summarization of information content of the taxon in a nested framework. A systemofnamingplantsforscientificcommunicationmustbe international inscope,andmustprovideconsistencyintheapplicationof names.Itmustalsobeacceptedbymost,ifnotall,membersofthe scientific community. These criteria led, almost inevitably, to International Botanical Congresses (IBCs) being the venue at which agreement on a system of scientific nomenclature for plants was sought. The IBCs led to publication of different ‘Codes’ which embodied the rules and regulations of botanical nomenclature and the decisions taken during these Congresses. Advantages ofBotanical Nomenclature: Though a common name may be much easier to remember, there are several good reasons to use botanical names for plant identification. Common names are not unique to a specific plant. -

Historical Review of Systematic Biology and Nomenclature - Alessandro Minelli
BIOLOGICAL SCIENCE FUNDAMENTALS AND SYSTEMATICS – Vol. II - Historical Review of Systematic Biology and Nomenclature - Alessandro Minelli HISTORICAL REVIEW OF SYSTEMATIC BIOLOGY AND NOMENCLATURE Alessandro Minelli Department of Biology, Via U. Bassi 58B, I-35131, Padova,Italy Keywords: Aristotle, Belon, Cesalpino, Ray, Linnaeus, Owen, Lamarck, Darwin, von Baer, Haeckel, Sokal, Sneath, Hennig, Mayr, Simpson, species, taxa, phylogeny, phenetic school, phylogenetic school, cladistics, evolutionary school, nomenclature, natural history museums. Contents 1. The Origins 2. From Classical Antiquity to the Renaissance Encyclopedias 3. From the First Monographers to Linnaeus 4. Concepts and Definitions: Species, Homology, Analogy 5. The Impact of Evolutionary Theory 6. The Last Few Decades 7. Nomenclature 8. Natural History Collections Glossary Bibliography Biographical Sketch Summary The oldest roots of biological systematics are found in folk taxonomies, which are nearly universally developed by humankind to cope with the diversity of the living world. The logical background to the first modern attempts to rationalize the classifications was provided by Aristotle's logic, as embodied in Cesalpino's 16th century classification of plants. Major advances were provided in the following century by Ray, who paved the way for the work of Linnaeus, the author of standard treatises still regarded as the starting point of modern classification and nomenclature. Important conceptual progress was due to the French comparative anatomists of the early 19th century UNESCO(Cuvier, Geoffroy Saint-Hilaire) – andEOLSS to the first work in comparative embryology of von Baer. Biological systematics, however, was still searching for a unifying principle that could provide the foundation for a natural, rather than conventional, classification.SAMPLE This principle wasCHAPTERS provided by evolutionary theory: its effects on classification are already present in Lamarck, but their full deployment only happened in the 20th century. -

Rules of Botanical Nomenclature
Taxonomy Prof.(Dr.) Punam Jeswal Head M.Sc semester II Botany Department Rules of Botanical Nomenclature Definition - Nomenclature is the art of naming of objects, which deals with the determination of a correct name to a known plant or to a known taxon. The names indeed correspond to the sentence, as both constitute meaningful collection of words. A name indicates a noun that helps in the quick identification, easy communication and economy of memory about the object to which it is concerned. Types of Names - The names according to their range of audience, language, territorial coverage and governance are of two types :- 1. Common or vernacular names. 2. International or scientific names. Common or Vernacular Names - These names are of the locals, by the locals, for the locals, in the local dialect. That is when a local plant is named by native people for the identification and communication to the other people of the same territory in their won local dialect, it is referred to as local or common or vernacular name. The fundamental demerits of this name are that they have limited audience, small territorial coverage and not governed under any set of principles or rules and even the same plant may have more than one name in the same locality. Another demerit of concern regarding these names is presence of synonyms in the languages therefore; the same plant may have a variety of names at different places in different languages. As for instance, mango(Mangifera indica) posses more than fifty names in Sanskrit only and lotus is known by more than two dozen names in Sanskrit and Hindi languages. -

Biological Nomenclature Free Download Pdf Book Charles Jeffrey
Biological Nomenclature Charles Jeffrey; Systematics Association Guidelines on Biological Nomenclature - Department of the . But have you ever wondered how all these organisms are named? In this lesson, you'll learn about the scientific naming system called binomial nomenclature. Binomial nomenclature - Wikipedia, the free encyclopedia Binomial nomenclature - New World Encyclopedia Biological nomenclature terms for facilitating communication in the . Scientific nomenclature was proposed by the Swedish biologist Carl von Linné . In current phylogenetic biological classification, organisms are grouped into BIOLOGICAL NOMENCLATURE and TAXONOMY DATA STANDARD terminology; a classified system of technical names, such as of anatomical structures or organisms. binomial nomenclature the nomenclature used in scientific Are the Linnean and Phylogenetic Nomenclatural Systems . 10 Sep 2009 . Carolus Linnaeus popularized the use of the binomial nomenclature within In biology, binomial nomenclature is the formal system of naming Binomial Nomenclature: Definition, Classification & System - Video . 8 May 2012 . A set of terms recommended for use in facilitating communication in biological nomenclature is presented as a table showing broadly Summary of the Linnean system of binomial nomenclature, the scientific way to name living things with a generic (genus) and specific (species) name. ENY-731/IN661: Scientific Nomenclature: What's in a Name? - EDIS a system of nomenclature in which each species of animal or plant receives a name of two terms of which the first identifies the genus to which it belongs and the . Scientific Classification - Fact Monster As will be seen below the various Codes for nomenclature consider Latin to be an essential language. Taxa at the level of species are named with binomials, consisting of generic and specific epithets or names that together equal the species name. -

Binomial Nomenclature B.Sc. Botany Sub. II Group
Dr. Sanjeev Kumar Vidyarthi Department of Botany Dr. L.K.V.D. College, Tajpur, Samastipur L.N. Mithila University, Darbhanga Botanical nomenclature The system currently used in applying names to plants, known as nomenclature, had its beginning with Carolus Linneaus. Species names have three components: (1) the genus name; (2) the specific epithet; and (3) the authority or individual(s) responsible for the name. Components 1 and 2 are either italicized or underlined. An example is Quercus alba L. Quercus is the genus name for the group of plants commonly known as oaks. The specific epithet is alba, Latin for white, and is descriptive of the bark and wood of the plant commonly known as white oak. The authority is L., an abbreviation for Linneaus, who first coined a formal name for this plant. Since the time of Linnaeus, the system of nomenclature has become more formalized and codified. The International Code of Botanical Nomenclature (ICBN) has been established to provide a uniform set of rules to be followed in applying names to plants. The rules contained in the ICBN are revised during the International Botanical Congresses, which are held every six years. Carolus Linneaus The core of the ICBN is composed of six principles- 1. "Botanical nomenclature is independent of zoological nomenclature." The rules of the ICBN do not apply to animals and bacteria. Therefore botanists do not have to be concerned with the names or rules associated with animals and bacteria. 2. "The application of names of taxonomic groups is determined by means of nomenclatural types." Each plant has a physical (type) specimen associated with it. -
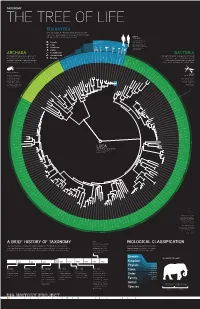
A Brief History of Taxonomy Biological Classification
TAXONOMY THE TREE OF LIFE EUKARYOTA This domain includes all of the plants, animals, and fungi, and some single-celled organisms. Eukaryotes are distinguished by their complex cells, which contain a membrane-enclosed nucleus. Humans Homo sapiens The creatures most familiar to us, Our species, primates in the animals, are members of the Animalia kingdom of the Animalia same kingdom. Eukaryota, is thought to have Fungi Mosquito Red first evolved in Africa about Pufferfish Junglefowl Roundworm Mouse 200,000 years ago. Genetically, Amoebozoa Chimpanzee our closest living relative Plantae is the chimpanzee. Archaeplastida Schizosaccharomyces pombe ARCHAEA Saccharomyces cerevisiae BACTERIA Caenorhabditis briggsae Caenorhabditis elegans Eremothecium gossypii These single-celled prokaryotic organisms often Chromalveolata Dictyostelium discoideum These single-celled prokaryotic organisms were among Cyanidioschyzon merolae live in extreme environmental conditions. Once Excavata Arabidopsis thaliana the first life forms to appear on Earth. Often spherical, Plasmodium falciparum considered to be Bacteria, these microorganisms Cryptosporidium hominis rod-like, or spiral in shape, these microorganisms Thalassiosira pseudonana Oryza sativa Anopheles gambiae Drosophila melanogaster Takifugu rubripes Danio rerio are now recognized as a separate domain of life. Gallus gallus function without a membrane-enclosed cell nucleus. Rattus norvegicus Mus musculus Methanococcus jannaschii Leishmania major Homo sapiens Pan troglodytes Methanococcus maripaludi Thermoanaerobacter -

Biblioqraphy & Natural History
BIBLIOQRAPHY & NATURAL HISTORY Essays presented at a Conference convened in June 1964 by Thomas R. Buckman Lawrence, Kansas 1966 University of Kansas Libraries University of Kansas Publications Library Series, 27 Copyright 1966 by the University of Kansas Libraries Library of Congress Catalog Card number: 66-64215 Printed in Lawrence, Kansas, U.S.A., by the University of Kansas Printing Service. Introduction The purpose of this group of essays and formal papers is to focus attention on some aspects of bibliography in the service of natural history, and possibly to stimulate further studies which may be of mutual usefulness to biologists and historians of science, and also to librarians and museum curators. Bibli• ography is interpreted rather broadly to include botanical illustration. Further, the intent and style of the contributions reflects the occasion—a meeting of bookmen, scientists and scholars assembled not only to discuss specific examples of the uses of books and manuscripts in the natural sciences, but also to consider some other related matters in a spirit of wit and congeniality. Thus we hope in this volume, as in the conference itself, both to inform and to please. When Edwin Wolf, 2nd, Librarian of the Library Company of Phila• delphia, and then Chairman of the Rare Books Section of the Association of College and Research Libraries, asked me to plan the Section's program for its session in Lawrence, June 25-27, 1964, we agreed immediately on a theme. With few exceptions, we noted, the bibliography of natural history has received little attention in this country, and yet it is indispensable to many biologists and to historians of the natural sciences. -

Binomial Nomenclature and Classification
FACULTY OF AGRICULTURAL SCIENCES AND ALLIED INDUSTRIES INTRODUCTORY BIOLOGY UGR-121 Binomial nomenclature and Classification LECTURE- 03 NOMENCLATURE Nomenclature is giving distinct scientific names to various structures including living organisms for their identification. The names are of two types - vernacular (common name) and scientific names. The vernacular names are based on some peculiarity of the organisms, e.g., Kandali (a plant having spines). Scientific names are distinct and specific, they have particular spellings which are not changed. TYPES OF NOMENCLATURE Three types of nomenclature are polynomial, binomial and trinomial. POLYNOMIAL SYSTEM According to this system, name of any plant consists of many words. For e.g., Caryophyllum saxatilis folis gramineus umbellatus corymbia which means Caryophyllum growing on rocks, having grass like leaves and umbellate corymb flowers. BINOMIAL SYSTEM Carolus Linnaeus used this nomenclature system for the first time and proposed scientific name of all the plants and animals. He is the founder of binomial system. Linnaeus proposed scientific name of plants in his book "Species plantarum". In binomial nomenclature, each scientific name has 2 components - generic name (genus) and specific name (species). E.g., Solanum tuberosum (potato), Mangifera indica (mango) The name indicates relationship of a species with others present in the same genus. TRINOMIAL SYSTEM This system was proposed by Huxley and Stricklandt. According to this system, name of any plant or species is composed of three names- Subspecific name (Name of variety) When members of any species have large variations then trinomial system is used. On the basis of Generic name Specific name dissimilarities, this species is classified into sub-species. -

History of Taxonomy
History of Taxonomy The history of taxonomy dates back to the origin of human language. Western scientific taxonomy started in Greek some hundred years BC and are here divided into prelinnaean and postlinnaean. The most important works are cited and the progress of taxonomy (with the focus on botanical taxonomy) are described up to the era of the Swedish botanist Carl Linnaeus, who founded modern taxonomy. The development after Linnaeus is characterized by a taxonomy that increasingly have come to reflect the paradigm of evolution. The used characters have extended from morphological to molecular. Nomenclatural rules have developed strongly during the 19th and 20th century, and during the last decade traditional nomenclature has been challenged by advocates of the Phylocode. Mariette Manktelow Dept of Systematic Biology Evolutionary Biology Centre Uppsala University Norbyv. 18D SE-752 36 Uppsala E-mail: [email protected] 1. Pre-Linnaean taxonomy 1.1. Earliest taxonomy Taxonomy is as old as the language skill of mankind. It has always been essential to know the names of edible as well as poisonous plants in order to communicate acquired experiences to other members of the family and the tribe. Since my profession is that of a systematic botanist, I will focus my lecture on botanical taxonomy. A taxonomist should be aware of that apart from scientific taxonomy there is and has always been folk taxonomy, which is of great importance in, for example, ethnobiological studies. When we speak about ancient taxonomy we usually mean the history in the Western world, starting with Romans and Greek. However, the earliest traces are not from the West, but from the East. -

Classification
Classification Say Thanks to the Authors Click http://www.ck12.org/saythanks (No sign in required) To access a customizable version of this book, as well as other interactive content, visit www.ck12.org CK-12 Foundation is a non-profit organization with a mission to reduce the cost of textbook materials for the K-12 market both in the U.S. and worldwide. Using an open-content, web-based collaborative model termed the FlexBook®, CK-12 intends to pioneer the generation and distribution of high-quality educational content that will serve both as core text as well as provide an adaptive environment for learning, powered through the FlexBook Platform®. Copyright © 2013 CK-12 Foundation, www.ck12.org The names “CK-12” and “CK12” and associated logos and the terms “FlexBook®” and “FlexBook Platform®” (collectively “CK-12 Marks”) are trademarks and service marks of CK-12 Foundation and are protected by federal, state, and international laws. Any form of reproduction of this book in any format or medium, in whole or in sections must include the referral attribution link http://www.ck12.org/saythanks (placed in a visible location) in addition to the following terms. Except as otherwise noted, all CK-12 Content (including CK-12 Curriculum Material) is made available to Users in accordance with the Creative Commons Attribution/Non- Commercial/Share Alike 3.0 Unported (CC BY-NC-SA) License (http://creativecommons.org/licenses/by-nc-sa/3.0/), as amended and updated by Creative Commons from time to time (the “CC License”), which is incorporated herein by this reference. -

Unit 4 Binomial Nomenclature
UNIT 4 BINOMIAL NOMENCLATURE Structure 4.1 Introduction Objectives 4.2 Development of Concepts International Codes Principles of Bionomial Nomenclature 4.3 Important Rules of Nomenclature 4.4 Binomial System Bauhin Linnaeus 4.5 Units of Classification Species, Genera and Families 4.6 Summary 4.7 Terminal Questions 4.8 Answers : 4.1 INTRODUCTION In Units 2 and 3 you have read about classification of plants, animals, the process of establishing and defining systematic grouping. In this unit we will discuss Birlqnial Nomenclature which is unique in Biological Sciences. Binomial nomenclature is the naming of the individual entity and the group so produced through classification, according to the international rules. These rules are publishdd in the form of international codes, Like all other branchei of science the field of biology is ever changing in all its contents like characters, circumscriptions and knowledge. Under the heading of binomial nomenclature we will discuss (1) its concepts and principles, (2) binomial systems as put forth by Bauhin and Linnaeus and (3) units of classification like species, genera and families..It is important to mention here that the international codes have no hand in deciding the scientific interpretation but whichever decision you follow the code should guide as to which name or names are to be applied to the entities under study. Objectives After reading this unit you should be able to: 0 Explain the concepts of binomial nomenclature 0 Apply the principles of binomial nomenclature to the naming of plants and animals Justify the importance of scientific names @ enlist international codes of nomenclature.