Effects of Forest Edges on Ovenbird Demography in a Managed Forest Landscape
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Systematic Relationships and Biogeography of the Tracheophone Suboscines (Aves: Passeriformes)
MOLECULAR PHYLOGENETICS AND EVOLUTION Molecular Phylogenetics and Evolution 23 (2002) 499–512 www.academicpress.com Systematic relationships and biogeography of the tracheophone suboscines (Aves: Passeriformes) Martin Irestedt,a,b,* Jon Fjeldsaa,c Ulf S. Johansson,a,b and Per G.P. Ericsona a Department of Vertebrate Zoology and Molecular Systematics Laboratory, Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden b Department of Zoology, University of Stockholm, SE-106 91 Stockholm, Sweden c Zoological Museum, University of Copenhagen, Copenhagen, Denmark Received 29 August 2001; received in revised form 17 January 2002 Abstract Based on their highly specialized ‘‘tracheophone’’ syrinx, the avian families Furnariidae (ovenbirds), Dendrocolaptidae (woodcreepers), Formicariidae (ground antbirds), Thamnophilidae (typical antbirds), Rhinocryptidae (tapaculos), and Conop- ophagidae (gnateaters) have long been recognized to constitute a monophyletic group of suboscine passerines. However, the monophyly of these families have been contested and their interrelationships are poorly understood, and this constrains the pos- sibilities for interpreting adaptive tendencies in this very diverse group. In this study we present a higher-level phylogeny and classification for the tracheophone birds based on phylogenetic analyses of sequence data obtained from 32 ingroup taxa. Both mitochondrial (cytochrome b) and nuclear genes (c-myc, RAG-1, and myoglobin) have been sequenced, and more than 3000 bp were subjected to parsimony and maximum-likelihood analyses. The phylogenetic signals in the mitochondrial and nuclear genes were compared and found to be very similar. The results from the analysis of the combined dataset (all genes, but with transitions at third codon positions in the cytochrome b excluded) partly corroborate previous phylogenetic hypotheses, but several novel arrangements were also suggested. -
Count Summary Report Printout Date: 11/11/2016
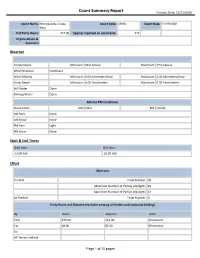
Count Summary Report Printout Date: 11/11/2016 Count Name: Monteverde, Costa Count Code: CRMO Count Date: 12/19/2001 Rica # of Party Hours: 257.00 Species reported on count date: 376 Organizations & Sponsors: Weather Temperature Minimum: 59.0 Celsius Maximum: 77.0 Celsius Wind Direction Northeast Wind Velocity Minimum: 0.00 Kilometers/hour Maximum: 6.00 Kilometers/hour Snow Depth Minimum: 0.00 Centimeters Maximum: 0.00 Centimeters Still Water Open Moving Water Open AM and PM Conditions Cloud Cover AM: Clear PM: Cloudy AM Rain None AM Snow None PM Rain Light PM Snow None Start & End Times Start time End time 12:00 AM 10:00 AM Effort Observers In Field Total Number: 70 Minimum Number of Parties (daylight): 19 Maximum Number of Parties (daylight): 24 At Feeders Total Number: 0 Party Hours and Distance (excludes viewing at feeders and nocturnal birding) By Hours Distance Units Foot 239.00 114.00 Kilometers Car 18.00 85.00 Kilometers Air All-Terrain Vehicle Page 1 of 12 pages Count Summary Report Printout Date: 11/11/2016 Bicycle Dog Sled Golfcart Horseback Motorized Boat Non-Motorized Boat Skis/Xc-Skis Snowmachine Snowshoe Wheelchair Other Time and Distance Hours Distance Units At Feeders 0.00 Nocturnal Birding 17.00 26.00 Miles Total Party 257.00 199.00 Kilometers Checklist Species Number Number/Party Hrs. Flags Editorial Codes Highland Tinamou 3 0.0117 Great Tinamou 2 0.0078 Gray-headed Chachalaca 33 0.1284 Crested Guan 20 0.0778 Black Guan 18 0.0700 Black-breasted Wood-Quail 56 0.2179 Least Grebe 5 0.0195 Anhinga 1 0.0039 Fasciated Tiger-Heron -

Ecology, Morphology, and Behavior in the New World Wood Warblers
Ecology, Morphology, and Behavior in the New World Wood Warblers A dissertation presented to the faculty of the College of Arts and Sciences of Ohio University In partial fulfillment of the requirements for the degree Doctor of Philosophy Brandan L. Gray August 2019 © 2019 Brandan L. Gray. All Rights Reserved. 2 This dissertation titled Ecology, Morphology, and Behavior in the New World Wood Warblers by BRANDAN L. GRAY has been approved for the Department of Biological Sciences and the College of Arts and Sciences by Donald B. Miles Professor of Biological Sciences Florenz Plassmann Dean, College of Arts and Sciences 3 ABSTRACT GRAY, BRANDAN L., Ph.D., August 2019, Biological Sciences Ecology, Morphology, and Behavior in the New World Wood Warblers Director of Dissertation: Donald B. Miles In a rapidly changing world, species are faced with habitat alteration, changing climate and weather patterns, changing community interactions, novel resources, novel dangers, and a host of other natural and anthropogenic challenges. Conservationists endeavor to understand how changing ecology will impact local populations and local communities so efforts and funds can be allocated to those taxa/ecosystems exhibiting the greatest need. Ecological morphological and functional morphological research form the foundation of our understanding of selection-driven morphological evolution. Studies which identify and describe ecomorphological or functional morphological relationships will improve our fundamental understanding of how taxa respond to ecological selective pressures and will improve our ability to identify and conserve those aspects of nature unable to cope with rapid change. The New World wood warblers (family Parulidae) exhibit extensive taxonomic, behavioral, ecological, and morphological variation. -

Passerines: Perching Birds
3.9 Orders 9: Passerines – perching birds - Atlas of Birds uncorrected proofs 3.9 Atlas of Birds - Uncorrected proofs Copyrighted Material Passerines: Perching Birds he Passeriformes is by far the largest order of birds, comprising close to 6,000 P Size of order Cardinal virtues Insect-eating voyager Multi-purpose passerine Tspecies. Known loosely as “perching birds”, its members differ from other Number of species in order The Northern or Common Cardinal (Cardinalis cardinalis) The Common Redstart (Phoenicurus phoenicurus) was The Common Magpie (Pica pica) belongs to the crow family orders in various fine anatomical details, and are themselves divided into suborders. Percentage of total bird species belongs to the cardinal family (Cardinalidae) of passerines. once thought to be a member of the thrush family (Corvidae), which includes many of the larger passerines. In simple terms, however, and with a few exceptions, passerines can be described Like the various tanagers, grosbeaks and other members (Turdidae), but is now known to belong to the Old World Like many crows, it is a generalist, with a robust bill adapted of this diverse group, it has a thick, strong bill adapted to flycatchers (Muscicapidae). Its narrow bill is adapted to to feeding on anything from small animals to eggs, carrion, as small birds that sing. feeding on seeds and fruit. Males, from whose vivid red eating insects, and like many insect-eaters that breed in insects, and grain. Crows are among the most intelligent of The word passerine derives from the Latin passer, for sparrow, and indeed a sparrow plumage the family is named, are much more colourful northern Europe and Asia, this species migrates to Sub- birds, and this species is the only non-mammal ever to have is a typical passerine. -

MS1001 Greeney
The nest of the Pearled Treerunner (Margarornis squamiger ) Artículo El nido del Subepalo Perlado (Margarornis squamiger) Harold F. Greeney1 & Rudy A. Gelis2 1Yanayacu Biological Station & Center for Creative Studies, Cosanga, Napo Province, Ecuador, c/o 721 Foch y Amazonas, Quito, Ecuador. [email protected] 2Pluma Verde Tours, Pasaje Manuel Garcia y 18 de Septiembre N20-28 Quito, Ecuador. Abstract Ornitología Colombiana Ornitología The Pearled Treerunner (Margarornis squamiger) is a small ovenbird (Furnariidae) inhabiting the upper strata of Neotropical montane forests. Little is known of its breeding habits despite its wide distribution and abundance within appropriate habitat. The genus Margarornis is considered closely related to Premnoplex barbtails, but details of nest architecture supporting this relationship are unavailable. Here we provide the first detailed description of nest architecture for the Pearled Treerunner from a nest encountered in northwest Ecuador. The nest was a tightly woven ball of moss and rootlets, similar in shape to that of the Spotted Barbtail (Premnoplex brunnescens) and presumably built in a similar manner. Nest architecture and nes- tling behavior support a close relationship between Margarornis and Premnoplex. Key words: barbtail, Ecuador, Furnariidae, Margarornis squamiger, nest architecture. Resumen El Subepalo Perlado (Margarornis squamiger) es un furnárido pequeño que habita el dosel de los bosques de montaña neo- tropicales. La reproducción de esta especie es poco conocida, a pesar de su amplia distribución y de ser común en el hábi- tat adecuado. El género Margarornis se considera cercanamente emparentado con los subepalos del género Premnoplex, pero no se conocen detalles de la arquitectura del nido que sustenten esta relación. -

Wood Warblers Wildlife Note
hooded warbler 47. Wood Warblers Like jewels strewn through the woods, Pennsylvania’s native warblers appear in early spring, the males arrayed in gleaming colors. Twenty-seven warbler species breed commonly in Pennsylvania, another four are rare breeders, and seven migrate through Penn’s Woods headed for breeding grounds farther north. In central Pennsylvania, the first species begin arriving in late March and early April. Louisiana waterthrush (Parkesia motacilla) and black and white warbler (Mniotilta varia) are among the earliest. The great mass of warblers passes through around mid-May, and then the migration trickles off until it ends in late May by which time the trees have leafed out, making it tough to spot canopy-dwelling species. In southern Pennsylvania, look for the migration to begin and end a few days to a week earlier; in northern Pennsylvania, it is somewhat later. As summer progresses and males stop singing on territory, warblers appear less often, making the onset of fall migration difficult to detect. Some species begin moving south as early as mid and late July. In August the majority specific habitat types and show a preference for specific of warblers start moving south again, with migration characteristics within a breeding habitat. They forage from peaking in September and ending in October, although ground level to the treetops and eat mainly small insects stragglers may still come through into November. But by and insect larvae plus a few fruits; some warblers take now most species have molted into cryptic shades of olive flower nectar. When several species inhabit the same area, and brown: the “confusing fall warblers” of field guides. -

Seiurus Aurocapilla) on VACA KEY, FLORIDA
Florida Field Naturalist 41(4):123-125, 2013. ECTOPARASITES COLLECTED FROM THE OVENBIRD (Seiurus aurocapilla) ON VACA KEY, FLORIDA LAWRENCE J. HRIBAR Florida Keys Mosquito Control District, 503 107th Street, Marathon, Florida 33050 The quill mite, Syringophiloidus seiurus (Clark) (Prostigmata: Syringophilidae) and the louse flyOrnithoctona fusciventris (Wiedemann) (Diptera: Hippoboscidae) are among the very few records of ectoparasites from the Ovenbird, Seiurus aurocapilla from Florida (Forrester and Spaulding 2003). On the 17th of November 2011, an Ovenbird was found dead outside a building on Vaca Key in Marathon, Florida (24.729984, -81.039438), apparently having collided with a plate glass window. The bird was handled and feather mites recovered and prepared for study in the same manner as were the specimens examined by Hribar and Miller (2011). Only twenty-five feather mites were recovered. Slide mounts were examined via phase contrast microscopy and then sent to a specialist for identification. Three mite species were recovered, two Proctophyllodidae (Proctophyllodes sp., Amerodectes sp.) and one Trouessartiidae. Unfortunately no specimens were readily identifiable to species. One female mite was identified as Proctophyllodes sp. Females of this genus are very difficult to identify to species, however,Proctophyllodes breviquadratus Atyeo and Braasch is known from Ovenbirds (Atyeo and Braasch 1966). One male and three female Amerodectes were not identifiable to species and may represent an undescribed species. Amerodectes mites are found on a variety of birds in the New World, viz., Apodiformes: Apodidae; Passeriformes: Cardinalidae, Emberizidae, Furnariidae, Hirundinidae, Icteridae, Parulidae, Thraupidae,and Turdidae (Valim and Hernandes 2010). The two male and two female Trouessartia mites appear to be conspecific with mites found on Ovenbirds in Alberta, Canada, and also represent an undescribed species (H. -

Lapa Rios Bird Checklist Lapa Rios Bird Checklist
Lapa Rios Bird Checklist Lapa Rios Bird Checklist The birds listed as "have been seen" at Lapa Rios include the Reserve itself as well as sighthings in the Matapalo (beach) area, in and around Puerto Jiménez and along the road from Puerto Jiménez to Lapa Rios; a distance of approximately 19 kilometers (11 miles). Lapa Rios is a private Biological Preserve of approximately 1000 acres. Access to its trail system is only through the permission of the management. The trail inmmediately adjacent to the main lodge can be explored without a staff guide, but a staff guide is required for any excursion into the interior of the preserve or along the Carbonera River. STATUS CODE: A = "Abundant" - many seen or heard daily in appropriate habitat/season and/or in large groups at frequent intervals. C = "Common" - consistently recorded in appropiate habitat/season and/or in large groups at frequent intervals. U = "Uncommon" - recorded regularly but with longer intervals and in small numbers. R = "Rare" - recorded in very small numbers or on really rare occasions. Acc = "Accidental" - recorded only a few times at Lapa Rios sometimes far out of its normal range and not likely to recur. Ex = "Extinct"- considered to be extint in the wild, with no populations on the country and only few sightings in the last years. GARRIGUES GUIDE: We reference Richard Garrigues guidebook for the bird’s description. The Birds of Costa Rica: A Field Guide. Zona Tropical Publications, Paperback – April 12, 2007 1 COMMON NAME LATIN NAME STATUS GUIDE TINAMOUS 1 Great Tinamou Tinamus major A Pag. -

Great Rivers of the Amazon III: Mamirauá, Amanã, and Tefé 2019
Field Guides Tour Report Great Rivers of the Amazon III: Mamirauá, Amanã, and Tefé 2019 Dec 2, 2019 to Dec 16, 2019 Bret Whitney & Micah Riegner For our tour description, itinerary, past triplists, dates, fees, and more, please VISIT OUR TOUR PAGE. The White Uakari alone is a reason to visit this part of Amazonia. With its striking red face, purple eyeshadow and fluffy white fur, it's gotta be one of the most outlandish of all Neotropical primates. We had an amazing time watching a troop right above us at Mamiraua. Photo by Micah Riegner. Establishing a new tour in Amazonia where no birders go is always thrilling—you just never know what you’re going to find up the next igarape (creek), what the next feeding flock will bring, or night outing might yield. So, in perpetuating the Field Guides tradition of going to new exciting places, Bret and I visited Reserva Amana and FLONA Tefe back in June. These two enormous swaths of protected forest in the heart of Amazonia had never been visited by a tour group, so we had some ground-truthing to do before the tour. This involved spending hours in a voadeira (open-top metal boat with outboard motor) to reach the isolated communities within the reserves, meeting with the local people and explaining what bird-watching is all about and checking out the trails they had cleared in anticipation of our arrival. Our scouting was fruitful, and we are pleased to announce a successful first run of the tour! There were numerous highlights, from seeing between 1,500 and 2,000 Sand-colored Nighthawks dusting the treetops like snow at Amana to watching Margays almost getting into a cat fight in the canopy at Mamiraua and the band of White Uakaris with their intimidating red faces. -

Display Behavior of Ovenbirds (Seiurus Aurocapillus) I
W&m Bull., 92(3), 1980, pp. 312-329 DISPLAY BEHAVIOR OF OVENBIRDS (SEIURUS AUROCAPILLUS) I. NON-SONG VOCALIZATIONS M. Ross LEIN Although detailed investigations of single displays have furnished in- sights into their proximate causation and adaptive roles in regulating social behavior (Smith 1977, Sebeok 1977), they contribute relatively little to our understanding of the evolution of animal communication at a somewhat higher level. Smith (1969a) and Moynihan (1970) pioneered consideration of the evolutionary pressures shaping the total display repertoire of a species. However, such studies are hampered by a paucity of information about the size and composition of repertoires of most animals, since de- tailed inventories are time-consuming and are frequently by-passed by workers studying single displays or small portions of a repertoire. Gen- eralizations are therefore based on a few species whose display behavior is known in sufficient detail, and their validity depends on how well ad- ditional studies conform to the patterns found. During investigations of behavior of wood warblers (Parulidae) I ob- tained information on the display repertoires of various species. In this paper I describe the non-song vocalizations of breeding Ovenbirds (Seiurus aurocupillus) and suggest some of the functional aspects of vocal com- munication and the evolutionary pressures that possibly have shaped its display repertoire. The songs of males and visual displays will be dealt with in subsequent papers. The Ovenbird is an excellent subject for behavioral investigation. It is widely distributed and abundant throughout the wooded regions of North America, occupying open woodlands, generally with little underbrush but with an abundance of fallen leaves, logs and rocks (Bent 1953, Griscom and Sprunt 1957). -

Biosurvey News the Newsletter of the Oklahoma Biological Survey Summer 2007
Biosurvey News The Newsletter of the Oklahoma Biological Survey Summer 2007 Biological Survey to Lead Migration Network All summer long that scissor-tailed flycatcher sits to accelerate the flow of technology for use in track- on the phone line or fence in your neighborhood. ing migrants and to train the next generation of sci- Have you every wondered where “your flycatcher” entists in the latest tracking techniques. and all of Oklahoma’s other migrants spend their Some of the recent advances in tracking tech- winter months? A group of scientists has, and now nologies include: development of satellite transmit- the National Science Foundation has funded this ters that allow birds to be tracked continuously group so that they can try to find out. The across continents; molecular genetic and stable iso- Oklahoma Biological Survey is coordinating the tope analyses of tissues that allow breeding and Migration Interest Group: Research Applied Toward over-wintering localities to be identified and popula- Education (MIGRATE) network tions to be connected; and use of harmonic radar to (www.migrate.ou.edu). This network of scientists, track flights of individual invertebrates. The headed by Dr. Jeff Kelly, is interested in improving MIGRATE network will work to accelerate this our ability to track migrants through their seasonal progress by making connections among researchers journeys that span the Americas. Animals of all with different areas of expertise and by training stu- kinds undergo seasonal migrations. Whether we are dents to search across disciplinary boundaries for interested in butterflies, sea turtles, lampreys or the key technologies and questions that will move songbirds, each of these migrants face common migration science forward. -

Phylogenetic Analysis of the Nest Architecture of Neotropical Ovenbirds (Furnariidae)
The Auk 116(4):891-911, 1999 PHYLOGENETIC ANALYSIS OF THE NEST ARCHITECTURE OF NEOTROPICAL OVENBIRDS (FURNARIIDAE) KRZYSZTOF ZYSKOWSKI • AND RICHARD O. PRUM NaturalHistory Museum and Department of Ecologyand Evolutionary Biology, University of Kansas,Lawrence, Kansas66045, USA ABSTRACT.--Wereviewed the tremendousarchitectural diversity of ovenbird(Furnari- idae) nestsbased on literature,museum collections, and new field observations.With few exceptions,furnariids exhibited low intraspecificvariation for the nestcharacters hypothe- sized,with the majorityof variationbeing hierarchicallydistributed among taxa. We hy- pothesizednest homologies for 168species in 41 genera(ca. 70% of all speciesand genera) and codedthem as 24 derivedcharacters. Forty-eight most-parsimonious trees (41 steps,CI = 0.98, RC = 0.97) resultedfrom a parsimonyanalysis of the equallyweighted characters using PAUP,with the Dendrocolaptidaeand Formicarioideaas successiveoutgroups. The strict-consensustopology based on thesetrees contained 15 cladesrepresenting both tra- ditionaltaxa and novelphylogenetic groupings. Comparisons with the outgroupsdemon- stratethat cavitynesting is plesiomorphicto the furnariids.In the two lineageswhere the primitivecavity nest has been lost, novel nest structures have evolved to enclosethe nest contents:the clayoven of Furnariusand the domedvegetative nest of the synallaxineclade. Althoughour phylogenetichypothesis should be consideredas a heuristicprediction to be testedsubsequently by additionalcharacter evidence, this first cladisticanalysis