33-Adrenoceptor-Mediated Relaxation Induced by Isoprenaline And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Properties and Units in Clinical Pharmacology and Toxicology
Pure Appl. Chem., Vol. 72, No. 3, pp. 479–552, 2000. © 2000 IUPAC INTERNATIONAL FEDERATION OF CLINICAL CHEMISTRY AND LABORATORY MEDICINE SCIENTIFIC DIVISION COMMITTEE ON NOMENCLATURE, PROPERTIES, AND UNITS (C-NPU)# and INTERNATIONAL UNION OF PURE AND APPLIED CHEMISTRY CHEMISTRY AND HUMAN HEALTH DIVISION CLINICAL CHEMISTRY SECTION COMMISSION ON NOMENCLATURE, PROPERTIES, AND UNITS (C-NPU)§ PROPERTIES AND UNITS IN THE CLINICAL LABORATORY SCIENCES PART XII. PROPERTIES AND UNITS IN CLINICAL PHARMACOLOGY AND TOXICOLOGY (Technical Report) (IFCC–IUPAC 1999) Prepared for publication by HENRIK OLESEN1, DAVID COWAN2, RAFAEL DE LA TORRE3 , IVAN BRUUNSHUUS1, MORTEN ROHDE1, and DESMOND KENNY4 1Office of Laboratory Informatics, Copenhagen University Hospital (Rigshospitalet), Copenhagen, Denmark; 2Drug Control Centre, London University, King’s College, London, UK; 3IMIM, Dr. Aiguader 80, Barcelona, Spain; 4Dept. of Clinical Biochemistry, Our Lady’s Hospital for Sick Children, Crumlin, Dublin 12, Ireland #§The combined Memberships of the Committee and the Commission (C-NPU) during the preparation of this report (1994–1996) were as follows: Chairman: H. Olesen (Denmark, 1989–1995); D. Kenny (Ireland, 1996); Members: X. Fuentes-Arderiu (Spain, 1991–1997); J. G. Hill (Canada, 1987–1997); D. Kenny (Ireland, 1994–1997); H. Olesen (Denmark, 1985–1995); P. L. Storring (UK, 1989–1995); P. Soares de Araujo (Brazil, 1994–1997); R. Dybkær (Denmark, 1996–1997); C. McDonald (USA, 1996–1997). Please forward comments to: H. Olesen, Office of Laboratory Informatics 76-6-1, Copenhagen University Hospital (Rigshospitalet), 9 Blegdamsvej, DK-2100 Copenhagen, Denmark. E-mail: [email protected] Republication or reproduction of this report or its storage and/or dissemination by electronic means is permitted without the need for formal IUPAC permission on condition that an acknowledgment, with full reference to the source, along with use of the copyright symbol ©, the name IUPAC, and the year of publication, are prominently visible. -
Guidance for the Format and Content of the Protocol of Non-Interventional
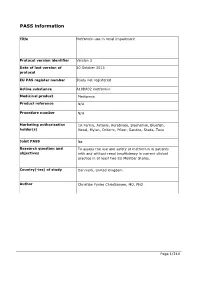
PASS information Title Metformin use in renal impairment Protocol version identifier Version 2 Date of last version of 30 October 2013 protocol EU PAS register number Study not registered Active substance A10BA02 metformin Medicinal product Metformin Product reference N/A Procedure number N/A Marketing authorisation 1A Farma, Actavis, Aurobindo, Biochemie, Bluefish, holder(s) Hexal, Mylan, Orifarm, Pfizer, Sandoz, Stada, Teva Joint PASS No Research question and To assess the use and safety of metformin in patients objectives with and without renal insufficiency in current clinical practice in at least two EU Member States. Country(-ies) of study Denmark, United Kingdom Author Christian Fynbo Christiansen, MD, PhD Page 1/214 Marketing authorisation holder(s) Marketing authorisation N/A holder(s) MAH contact person N/A Page 2/214 1. Table of Contents PASS information .......................................................................................................... 1 Marketing authorisation holder(s) .................................................................................... 2 1. Table of Contents ...................................................................................................... 3 2. List of abbreviations ................................................................................................... 4 3. Responsible parties .................................................................................................... 5 4. Abstract .................................................................................................................. -

Novel Method of Administering Β-Blockers and Novel Dosage Forms Containing Same Anwar A
University of Kentucky UKnowledge Pharmaceutical Sciences Faculty Patents Pharmaceutical Sciences 1-31-1984 Novel Method of Administering β-Blockers and Novel Dosage Forms Containing Same Anwar A. Hussain University of Kentucky Right click to open a feedback form in a new tab to let us know how this document benefits oy u. Follow this and additional works at: https://uknowledge.uky.edu/ps_patents Part of the Pharmacy and Pharmaceutical Sciences Commons Recommended Citation Hussain, Anwar A., "Novel Method of Administering β-Blockers and Novel Dosage Forms Containing Same" (1984). Pharmaceutical Sciences Faculty Patents. 124. https://uknowledge.uky.edu/ps_patents/124 This Patent is brought to you for free and open access by the Pharmaceutical Sciences at UKnowledge. It has been accepted for inclusion in Pharmaceutical Sciences Faculty Patents by an authorized administrator of UKnowledge. For more information, please contact [email protected]. Unlted States Patent [191 [11] 4,428,883 Hussain [45] Jan. 31, 1984 [54] NOVEL METHOD OF ADMINISTERING 4,012,444 3/l977 Lunts ................................. .. 424/ 324 B-BLOCKERSF0 RMS Co NTA' AND, INING NOVEL SAME DOSAGE ' 4,250,163, , 1.1’;2/l981 llieoll’loldNagaioe a ......................................... ---- -~.. .. 424/14 [7 5 ] I nventor : An war A . Hussam,° Lexington' , Ky . FOREIGN PATENT DOCUMENTS [73] Assignee: The University of Kentucky Research . Foundation’ Lexington, Ky. 979389 1/ 1965 United Klngdom . I [21] APPL Nod 241,413 OTHER PUBLICATIONS . Stern, Arzmit. Forsch, vol. 24, 1974, pp. 70-71. [22] Flled‘ Mm‘- 6’ 1981 Black, Brit. J. PharmacoL, (1965) 25, pp. 577-591. [51] Int. Cl.3 ................... .. A61K 27/00; A61K 31/15; Prima'y Examiner_stanley L Friedman A61K 31/40; A61K 31/47; A61K 31/135; Attorney, Agent, or Firm-Burns, Doane, Swecker & A61K 31/165; A61K 31/435; A61K 31/475; Mathis A61K 47/00; C07C 143/90; CllD 1/28; C09F - 5 /()() [57] ABSTRACT [52] US. -
![Ehealth DSI [Ehdsi V2.2.2-OR] Ehealth DSI – Master Value Set](https://docslib.b-cdn.net/cover/8870/ehealth-dsi-ehdsi-v2-2-2-or-ehealth-dsi-master-value-set-1028870.webp)
Ehealth DSI [Ehdsi V2.2.2-OR] Ehealth DSI – Master Value Set
MTC eHealth DSI [eHDSI v2.2.2-OR] eHealth DSI – Master Value Set Catalogue Responsible : eHDSI Solution Provider PublishDate : Wed Nov 08 16:16:10 CET 2017 © eHealth DSI eHDSI Solution Provider v2.2.2-OR Wed Nov 08 16:16:10 CET 2017 Page 1 of 490 MTC Table of Contents epSOSActiveIngredient 4 epSOSAdministrativeGender 148 epSOSAdverseEventType 149 epSOSAllergenNoDrugs 150 epSOSBloodGroup 155 epSOSBloodPressure 156 epSOSCodeNoMedication 157 epSOSCodeProb 158 epSOSConfidentiality 159 epSOSCountry 160 epSOSDisplayLabel 167 epSOSDocumentCode 170 epSOSDoseForm 171 epSOSHealthcareProfessionalRoles 184 epSOSIllnessesandDisorders 186 epSOSLanguage 448 epSOSMedicalDevices 458 epSOSNullFavor 461 epSOSPackage 462 © eHealth DSI eHDSI Solution Provider v2.2.2-OR Wed Nov 08 16:16:10 CET 2017 Page 2 of 490 MTC epSOSPersonalRelationship 464 epSOSPregnancyInformation 466 epSOSProcedures 467 epSOSReactionAllergy 470 epSOSResolutionOutcome 472 epSOSRoleClass 473 epSOSRouteofAdministration 474 epSOSSections 477 epSOSSeverity 478 epSOSSocialHistory 479 epSOSStatusCode 480 epSOSSubstitutionCode 481 epSOSTelecomAddress 482 epSOSTimingEvent 483 epSOSUnits 484 epSOSUnknownInformation 487 epSOSVaccine 488 © eHealth DSI eHDSI Solution Provider v2.2.2-OR Wed Nov 08 16:16:10 CET 2017 Page 3 of 490 MTC epSOSActiveIngredient epSOSActiveIngredient Value Set ID 1.3.6.1.4.1.12559.11.10.1.3.1.42.24 TRANSLATIONS Code System ID Code System Version Concept Code Description (FSN) 2.16.840.1.113883.6.73 2017-01 A ALIMENTARY TRACT AND METABOLISM 2.16.840.1.113883.6.73 2017-01 -

An Insight on Analytical Profile on Bisoprolol Fumarate – a Selective Beta-1 Adrenoreceptor Blocker
DOI: 10.15415/jptrm.2017.52012 An Insight on Analytical Profile on Bisoprolol Fumarate – A Selective Beta-1 Adrenoreceptor Blocker AJINKYA G. DHANDAR, SURAJ R. CHAUDHARI, SAURABH B. GANORKAR, AMOD S. PATIL, SANJAY J. SURANA AND ATUL A. SHIRKHEDKAR* R.C. Patel Institute of Pharmaceutical Education and Research, Shirpur, Dist. Dhule, India *Email: [email protected] Received: June 15, 2017 | Revised: July 19, 2017 | Accepted: Sept. 22, 2017 Published online: Nov. 02, 2017 The Author(s) 2017. This article is published with open access at www.chitkara.edu.in/publications Abstract BF is Beta-adreno receptor antagonist and used as an Anti- Hypertensive Drug. BF gives the blocking action on β1-adrenergic receptors in the heart and vascular smooth muscle. The present review compiles the various approaches implemented for quantification of BF in bulk drug, pharmaceutical matrix and biological fluid. This review represents more than 50 analytical methods which include capillary electrophoresis, HPLC, HPTLC, UV-Spectroscopy, UPLC, impurity profiling and electrochemical methods implemented for estimation of BF as a single component as well as in multicomponent. Keyword: BF; Bioanalytical; UPLC/LC-MS; capillary electrophoresis; impurity profile 1. INTRODUCTION BF is an extremely discriminatory β1-adrenergic blocker [1]. BF is chemically: (RS)-1-[4-[[2-(1- Methylethoxy) ethoxy] methyl] phenoxy] -3-[(1 methyl ethyl) amino] propan-2-ol fumarate Figure 1. It is official in, USP. BF has similar structure to metoprolol, bopindolol, hydrochlorothiazide, atenolol [2]. Structure of BF, there is two substituents β at para position of benzene provide the activity of -selectivity, In which Journal of Pharmaceutical it has two substituents in para position of benzene which might be the Technology, Research and activity of β- selectivity [3]. -

Drugs for Primary Prevention of Atherosclerotic Cardiovascular Disease: an Overview of Systematic Reviews
Supplementary Online Content Karmali KN, Lloyd-Jones DM, Berendsen MA, et al. Drugs for primary prevention of atherosclerotic cardiovascular disease: an overview of systematic reviews. JAMA Cardiol. Published online April 27, 2016. doi:10.1001/jamacardio.2016.0218. eAppendix 1. Search Documentation Details eAppendix 2. Background, Methods, and Results of Systematic Review of Combination Drug Therapy to Evaluate for Potential Interaction of Effects eAppendix 3. PRISMA Flow Charts for Each Drug Class and Detailed Systematic Review Characteristics and Summary of Included Systematic Reviews and Meta-analyses eAppendix 4. List of Excluded Studies and Reasons for Exclusion This supplementary material has been provided by the authors to give readers additional information about their work. © 2016 American Medical Association. All rights reserved. 1 Downloaded From: https://jamanetwork.com/ on 09/28/2021 eAppendix 1. Search Documentation Details. Database Organizing body Purpose Pros Cons Cochrane Cochrane Library in Database of all available -Curated by the Cochrane -Content is limited to Database of the United Kingdom systematic reviews and Collaboration reviews completed Systematic (UK) protocols published by by the Cochrane Reviews the Cochrane -Only systematic reviews Collaboration Collaboration and systematic review protocols Database of National Health Collection of structured -Curated by Centre for -Only provides Abstracts of Services (NHS) abstracts and Reviews and Dissemination structured abstracts Reviews of Centre for Reviews bibliographic -

Adrenoceptor Agonists on Spontaneous Contractions of Human Nonpregnant Myometrium
PRACE ORYGINALNE Ginekol Pol. 2011, 82, 918-924 ginekologia Differences in the effects ofβ 2- and β3- adrenoceptor agonists on spontaneous contractions of human nonpregnant myometrium Odmienny wpływ agonistów receptorów β2- i β3-adrenergicznych na spontaniczne skurcze myometrium kobiet nieciężarnych Pędzińska-Betiuk Anna1*, Modzelewska Beata1, Jóźwik Marcin2, Jóźwik Maciej3, Kostrzewska Anna1 1 Department of Biophysics, Medical University of Białystok, Białystok, Poland 2 Department of Gynecology and Obstetrics, Kliniken Nordoberpfalz, Akademisches Lehrkrankenhaus der Universität Regensburg, Weiden, Germany 3 Department of Gynecology, Medical University of Białystok, Białystok, Poland * Current address: Department of Experimental Physiology and Pathophysiology, Medical University of Białystok, Białystok, Poland Abstract Objective: This study aimed to compare the relaxant properties of BRL 37344 with β2-adrenoceptors agonist ritodrine on the contractility of human nonpregnant myometrium. Material and methods: The activity of myometrial strips mounted in an organ bath was recorded under isometric conditions using force transducers with digital output. Contractility before and after cumulative additions of both uterorelaxants and with preincubation with β-adrenoceptor antagonists bupranolol, propranolol, and butoxamine were studied. Results: Both BRL 37344 (10-10 – 10-4 mol/L) and ritodrine (10-10 – 10-5 mol/L) decreased the area under curve, or AUC, value (logIC50 -6.45 ± 0.18 and -8.71 ± 0.35, respectively), and the degree of inhibition of spontaneous contractile activity was similar (< 30%). However, BRL 37344 decreased the mean frequency of contractions, whereas ritodrine decreased the mean amplitude of contractions. The inhibition of contractions by BRL 37344 was partially antagonized by bupranolol and propranolol, but not with butoxamine. The inhibition by ritodrine was counteracted by all these antagonists. -

Carvedilol Inhibits EGF-Mediated JB6 P+ Colony Formation Through a Mechanism Independent of Adrenoceptors
RESEARCH ARTICLE Carvedilol inhibits EGF-mediated JB6 P+ colony formation through a mechanism independent of adrenoceptors 1¤a 1¤b 1 1¤c 2 Kristan H. Cleveland , Sherry Liang , Andy Chang , Kevin M. HuangID , Si Chen , 2 1 1 Lei Guo , Ying Huang *, Bradley T. AndresenID * 1 Department of Pharmaceutical Sciences, College of Pharmacy, Western University of Health Sciences, Pomona, California, United States of America, 2 Division of Biochemical Toxicology, National Center for Toxicological Research, US Food and Drug Administration, Jefferson, Arkansas, United States of America a1111111111 a1111111111 ¤a Current address: Department of Pharmacology and Toxicology, College of Pharmacy, University of a1111111111 Arizona, Tucson, Arizona, United States of America. a1111111111 ¤b Current address: Department of Medicinal Chemistry and Molecular Pharmacology, College of Pharmacy, a1111111111 Purdue University, West Lafayette, Indiana, United States of America. ¤c Current address: Department of Pharmaceutics and Pharmaceutical Chemistry, College of Pharmacy, The Ohio State University, Columbus, Ohio, United States of America. * [email protected] (YH); [email protected] (BTA) OPEN ACCESS Abstract Citation: Cleveland KH, Liang S, Chang A, Huang KM, Chen S, Guo L, et al. (2019) Carvedilol inhibits Carvedilol is reported to prevent cancers in humans and animal models. However, a molec- EGF-mediated JB6 P+ colony formation through a mechanism independent of adrenoceptors. PLoS ular mechanism has yet to be established, and the extent to which other β-blockers are che- ONE 14(5): e0217038. https://doi.org/10.1371/ mopreventive remains relatively unknown. A comparative pharmacological approach was journal.pone.0217038 utilized with the expectation that a mechanism of action could be devised. -

Pharmacological Properties of Beta-Adrenoceptor Blocking Drugs
Journal of Clinical and Basic Cardiology An Independent International Scientific Journal Journal of Clinical and Basic Cardiology 1998; 1 (1), 5-9 Pharmacological properties of beta-adrenoceptor blocking drugs Borchard U Homepage: www.kup.at/jcbc Online Data Base Search for Authors and Keywords Indexed in Chemical Abstracts EMBASE/Excerpta Medica Krause & Pachernegg GmbH · VERLAG für MEDIZIN und WIRTSCHAFT · A-3003 Gablitz/Austria REVIEWS β-blocking drugs J Clin Bas Cardiol 1998; 1: 5 Pharmacological properties of β-adrenoceptor blocking drugs U. Borchard β-adrenoceptor blocking drugs are widely used for the treat- Pharmacodynamic properties ment of cardiovascular diseases such as arterial hypertension, β β coronary heart disease and supraventricular and ventricular In many organs there is a coexistence of 1- and 2-receptors (Table 1). For example, in the normal human heart about 80% tachyarrhythmias. They may also be beneficial in the hyper- β β β kinetic heart syndrome, hypotensive circulatory disorders, of the -receptors are of the 1-subtype. In heart failure 1- receptors are down-regulated so that a relatively higher pro- portal hypertension, hyperthyroidism, tremour, migraine, β anxiety, psychosomatic disorders or glaucoma. In recent years portion of 2-receptors can be measured [3]. The physiological and therapeutic actions of a β-blocker depend on the actual even patients with heart failure have been successfully treated β β with β-blockers initially given at very low doses. density of 1- and/or 2-receptors in the different organs, on β A great number of β-adrenoceptor blocking drugs are now the affinity of the -blocker and on the local drug concen- available for clinical use which differ widely with respect to tration. -

A Study on Beta Blockers - a Brief Review Tv
IJRPC 2018, 8(4), 508-529 Namratha et al. ISSN: 22312781 INTERNATIONAL JOURNAL OF RESEARCH IN PHARMACY AND CHEMISTRY Available online at www.ijrpc.com Review Article A STUDY ON BETA BLOCKERS - A BRIEF REVIEW TV. Namratha*, KC. Chaluvaraju and AM. Anushree Department of Pharmaceutical Chemistry, Government College of Pharmacy, Bengaluru-560 027, Karnataka, India. ABSTRACT Beta blockers are agents or drugs which competitively inhibit the action of catecholamines at the beta adrenergic receptors, which are mainly used to treat variety of clinical conditions like angina, hypertension, asthma, COPD and arrhythmias. These drugs are also useful in several other therapeutic situations including shock, premature labor and opioid withdrawal, and as adjuncts to general anesthetics. These drugs produce their effect by interacting with the beta adrenergic receptors. In the present communication, an effort has been made to compile beta adrenergic receptors and the chemistry, discovery and development, classification and therapeutic applications of beta blockers. Keywords: Beta blockers, adrenergic receptors, catecholamines and aryloxypropanolamines. 1. INTRODUCTION Beta blockers were first developed by Sir 1.1 Adrenergic receptors James Black in 1962. The structures of The ability of a molecule to selectively these receptors have been studied by x- agonize or antagonize adrenergic ray crystallography.5 In the current article receptor made great advances in beta blockers are majorly focused with pharmacotherapeutics. The discovery of respect to their location, functions adrenergic receptors lead to major mediated, discovery and development, development of newer adrenergic SAR, classification, structures and agonists as well as antagonist.1 therapeutic applications. Adrenergic receptors are 7- transmembrane spanning receptors 1.2 Locations and agonistic action of beta which mediate both central and adrenergic receptors6 peripheral actions of the adrenergic The following are the various beta neurotransmitters. -

Quality Assurance of Drug Value Sets for Clinical Quality Measures
Eliciting the intension of medication value sets Nathan Bahr Oregon Health & Science University [email protected] Mentors: Rainer Winnenburg PhD, Olivier Bodenreider MD, PhD National Library of Medicine, National Institutes of Health, Bethesda Maryland, USA {rainer.winnenburg|olivier.bodenreider}@nih.gov 1 / 28 Motivation • Meaningful Use specifies criteria that must be met to be eligible for incentive payments under the American Reinvestment and Recovery Act (ARRA). • Clinical Quality Measures (CQMs) are tools used to measure the quality of care provided. – Providers and hospitals must report CQMs to fulfill MU2 requirements. • Value Sets are lists of standardized terms used in CQMs. • The Value Set Authority Center (VSAC) is the part of the NLM that is responsible for validating and delivering the value sets. • Assist in Quality Assurance of value sets – Some work have been done on disease value sets – We look specifically at the medication value sets 2 / 28 Investigating 183 medication value sets • Examples of value set names: – Statin – Methylxanthines – IV Antibiotics Used For Prophylaxis for Colon Surgery – Antibiotic Medications for Pharyngitis – ACE Inhibitor or ARB – ADHD Medications • Value sets contain lists of clinical drugs or ingredients – We normalize to RxNorm IN • Atenolol 100 MG Oral Tablet Atenolol • Lipitor Atorvastatin 3 / 28 Objective Value set: Misc. Antidepressants Monoamine Oxidase Inhibitors Bupropion Isocarboxazid Isocarboxazid Phenelzine Phenelzine Selegiline Selegiline Tranylcypromine Tranylcypromine rasagukube vilazodone Eliciting the intension for medication value sets to support quality assurance and curation task of clinical quality measures. – Drug classes have an explicit, agreed-upon definition. – Elicitation is performed by mapping the definition of the class onto the value set intension when the drug members are similar. -

Review of Existing Classification Efforts
Project No. TREN-05-FP6TR-S07.61320-518404-DRUID DRUID Driving under the Influence of Drugs, Alcohol and Medicines Integrated Project 1.6. Sustainable Development, Global Change and Ecosystem 1.6.2: Sustainable Surface Transport 6th Framework Programme Deliverable 4.1.1 Review of existing classification efforts Due date of deliverable: (15.01.2008) Actual submission date: (07.02.2008) Start date of project: 15.10.2006 Duration: 48 months Organisation name of lead contractor for this deliverable: UGent Revision 1.0 Project co-funded by the European Commission within the Sixth Framework Programme (2002-2006) Dissemination Level PU Public X PP Restricted to other programme participants (including the Commission Services) RE Restricted to a group specified by the consortium (including the Commission Services) CO Confidential, only for members of the consortium (including the Commission Services) Task 4.1 : Review of existing classification efforts Authors: Kristof Pil, Elke Raes, Thomas Van den Neste, An-Sofie Goessaert, Jolien Veramme, Alain Verstraete (Ghent University, Belgium) Partners: - F. Javier Alvarez (work package leader), M. Trinidad Gómez-Talegón, Inmaculada Fierro (University of Valladolid, Spain) - Monica Colas, Juan Carlos Gonzalez-Luque (DGT, Spain) - Han de Gier, Sylvia Hummel, Sholeh Mobaser (University of Groningen, the Netherlands) - Martina Albrecht, Michael Heiβing (Bundesanstalt für Straßenwesen, Germany) - Michel Mallaret, Charles Mercier-Guyon (University of Grenoble, Centre Regional de Pharmacovigilance, France) - Vassilis Papakostopoulos, Villy Portouli, Andriani Mousadakou (Centre for Research and Technology Hellas, Greece) DRUID 6th Framework Programme Deliverable D.4.1.1. Revision 1.0 Review of Existing Classification Efforts Page 2 of 127 Introduction DRUID work package 4 focusses on the classification and labeling of medicinal drugs according to their influence on driving performance.