Immunohematology Vol 21, #4
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Study of Blood Group (ABO and RH) Association and Secretor Status of ABH Substances in Patients with Systemic Lupus Erythematosus
IOSR Journal of Dental and Medical Sciences (IOSR-JDMS) e-ISSN: 2279-0853, p-ISSN: 2279-0861.Volume 18, Issue 5 Ser. 12 (May. 2019), PP 31-35 www.iosrjournals.org Study of Blood Group (ABO and RH) Association and Secretor Status of ABH Substances in Patients with Systemic Lupus Erythematosus Dr. Tashi Paleng1, Dr. A. Barindra Sharma2, Dr.Santa Naorem3, Dr. Laikangbam Dayalaxmi4, Dr. Eric Hilton Wanniang5, Dr. Eswara Moorthy G6 1Postgraduate trainee, 2Professor & Head, 3Professor, 4Senoir Resident, 5Postgraduate trainee, 6Postgraduate trainee 1,2,4,5,6Department of Transfusion Medicine, 3Department of Medicine, Regional Institute of Medical Sciences, Imphal-795004, Manipur, India Corresponding Author: Dr. Tashi Paleng Abstract: Individuals who secrete their blood group antigens in the body fluids are called secretors and individuals who do not secrete are called non-secretors. Non-secretors of blood group antigens have been found to be associated with many infectious and autoimmune diseases by many previous studies. The aims of our study were to find out any association between ABO blood group, RhD blood group and secretor status of ABH substances with the patients of Systemic Lupus Erythematosus. This Case Control study was carried out in the Department of Transfusion Medicine in collaboration with the Department of Medicine, Regional Institute of Medical Sciences, Imphal Manipur from September 2016 to August 2018. 75 confirmed SLE patients attending Rheumatology OPD, Department of Medicine and 75 healthy sex matched voluntary blood donors attending blood bank, Department of Tranfusion Medicine have been enrolled in the study. Their blood were typed for ABO and RhD groups using conventional tube technique and saliva were tested for secretor and non-secretor status using heamagglutination inhibition test. -

Unintentional Platelet Removal by Plasmapheresis
Journal of Clinical Apheresis 16:55–60 (2001) Unintentional Platelet Removal by Plasmapheresis Jedidiah J. Perdue,1 Linda K. Chandler,2 Sara K. Vesely,1 Deanna S. Duvall,2 Ronald O. Gilcher,2 James W. Smith,2 and James N. George1* 1Hematology-Oncology Section, Department of Medicine, University of Oklahoma Health Sciences Center, Oklahoma City, Oklahoma 2Oklahoma Blood Institute, Oklahoma City, Oklahoma Therapeutic plasmapheresis may remove platelets as well as plasma. Unintentional platelet loss, if not recognized, may lead to inappropriate patient assessment and treatment. A patient with thrombotic thrombocytopenic purpura- hemolytic uremic syndrome (TTP-HUS) is reported in whom persistent thrombocytopenia was interpreted as continuing active disease; thrombocytopenia resolved only after plasma exchange treatments were stopped. This observation prompted a systematic study of platelet loss with plasmapheresis. Data are reported on platelet loss during 432 apheresis procedures in 71 patients with six disease categories using three different instruments. Com- paring the first procedure recorded for each patient, there was a significant difference among instrument types ,than with the COBE Spectra (1.6% (21 ס P<0.001); platelet loss was greater with the Fresenius AS 104 (17.5%, N) .With all procedures, platelet loss ranged from 0 to 71% .(24 ס or the Haemonetics LN9000 (2.6%, N (26 ס N Among disease categories, platelet loss was greater in patients with dysproteinemias who were treated for hyper- viscosity symptoms. Absolute platelet loss with the first recorded apheresis procedure, in the 34 patients who had a normal platelet count before the procedure, was also greater with the AS 104 (2.23 × 1011 platelets) than with the Spectra (0.29 × 1011 platelets) or the LN9000 (0.37 × 1011 platelets). -

Platelet-Rich Plasmapheresis: a Meta-Analysis of Clinical Outcomes and Costs
THE jOURNAL OF EXTRA-CORPOREAL TECHNOLOGY Original Article Platelet-Rich Plasmapheresis: A Meta-Analysis of Clinical Outcomes and Costs Chris Brown Mahoney , PhD Industrial Relations Center, Carlson School of Management, University of Minnesota, Minneapolis, MN Keywords: platelet-rich plasmapheresis, sequestration, cardiopulmonary bypass, outcomes, economics, meta-analysis Presented at the American Society of Extra-Corporeal Technology 35th International Conference, April 3-6, 1997, Phoenix, Arizona ABSTRACT Platelet-rich plasmapheresis (PRP) just prior to cardiopulmonary bypass (CPB) surgery is used to improve post CPB hemostasis and to minimize the risks associated with exposure to allogeneic blood and its components. Meta-analysis examines evidence ofPRP's impact on clinical outcomes by integrating the results across published research studies. Data on clinical outcomes was collected from 20 pub lished studies. These outcomes, DRG payment rates, and current national average costs were used to examine the impact of PRP on costs. This study provides evidence that the use of PRP results in improved clinical outcomes when compared to the identical control groups not receiving PRP. These improved clinical out comes result in subsequent lower costs per patient in the PRP groups. All clinical outcomes analyzed were improved: blood product usage, length of stay, intensive care stay, time to extu bation, incidence of cardiovascular accident, and incidence of reoperation. The most striking differences occur in use of all blood products, particularly packed red blood cells. This study provides an example of how initial expenditure on technology used during CPB results in overall cost savings. Estimated cost savings range from $2,505.00 to $4,209.00. -

Journal of Blood Group Serology and Molecular Genetics Volume 34, Number 1, 2018 CONTENTS
Journal of Blood Group Serology and Molecular Genetics VOLUME 34, N UMBER 1, 2018 This issue of Immunohematology is supported by a contribution from Grifols Diagnostics Solutions, Inc. Dedicated to advancement and education in molecular and serologic immunohematology Immunohematology Journal of Blood Group Serology and Molecular Genetics Volume 34, Number 1, 2018 CONTENTS S EROLOGIC M ETHOD R EVIEW 1 Warm autoadsorption using ZZAP F.M. Tsimba-Chitsva, A. Caballero, and B. Svatora R EVIEW 4 Proceedings from the International Society of Blood Transfusion Working Party on Immunohaematology Workshop on the Clinical Significance of Red Blood Cell Alloantibodies, Friday, September 2, 2016, Dubai A brief overview of clinical significance of blood group antibodies M.J. Gandhi, D.M. Strong, B.I. Whitaker, and E. Petrisli C A S E R EPORT 7 Management of pregnancy sensitized with anti-Inb with monocyte monolayer assay and maternal blood donation R. Shree, K.K. Ma, L.S. Er and M. Delaney R EVIEW 11 Proceedings from the International Society of Blood Transfusion Working Party on Immunohaematology Workshop on the Clinical Significance of Red Blood Cell Alloantibodies, Friday, September 2, 2016, Dubai A review of in vitro methods to predict the clinical significance of red blood cell alloantibodies S.J. Nance S EROLOGIC M ETHOD R EVIEW 16 Recovery of autologous sickle cells by hypotonic wash E. Wilson, K. Kezeor, and M. Crosby TO THE E DITOR 19 The devil is in the details: retention of recipient group A type 5 years after a successful allogeneic bone marrow transplant from a group O donor L.L.W. -
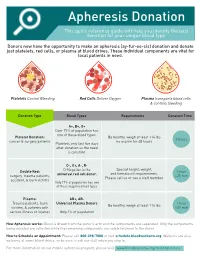
Apheresis Donation This Quick Reference Guide Will Help You Identify the Best Donation for Your Unique Blood Type
Apheresis Donation This quick reference guide will help you identify the best donation for your unique blood type. Donors now have the opportunity to make an apheresis (ay-fur-ee-sis) donation and donate just platelets, red cells, or plasma at blood drives. These individual components are vital for local patients in need. Platelets Control Bleeding Red Cells Deliver Oxygen Plasma transports blood cells & controls bleeding Donation Type Blood Types Requirements Donation Time A+, B+, O+ Over 75% of population has one of these blood types. Platelet Donation: Be healthy, weigh at least 114 lbs 2 hours cancer & surgery patients no aspirin for 48 hours Platelets only last five days after donation so the need is constant. O-, O+, A-, B- Special height, weight, Double Red: O-Negative is the 1 hour and hematocrit requirements. surgery, trauma patients, universal red cell donor. +25 min Please call us or see a staff member accident, & burn victims Only 17% of population has one of these negative blood types Plasma: AB+, AB- Trauma patients, burn Universal Plasma Donors 1 hour Be healthy, weigh at least 114 lbs victims, & patients with +30 min serious illness or injuries Only 4% of population How Apheresis works: Blood is drawn from the donor’s arm and the components are separated. Only the components being donated are collected while the remaining components are safely returned to the donor How to Schedule an Appointment: Please call 800-398-7888 or visit schedule.bloodworksnw.org. Walk-ins are also welcome at some blood drives, so be sure to ask our staff when you stop in. -

Blood Product Modifications: Leukofiltration, Irradiation and Washing
Blood Product Modifications: Leukofiltration, Irradiation and Washing 1. Leukocyte Reduction Definitions and Standards: o Process also known as leukoreduction, or leukofiltration o Applicable AABB Standards, 25th ed. Leukocyte-reduced RBCs At least 85% of original RBCs < 5 x 106 WBCs in 95% of units tested . Leukocyte-reduced Platelet Concentrates: At least 5.5 x 1010 platelets in 75% of units tested < 8.3 x 105 WBCs in 95% of units tested pH≥6.2 in at least 90% of units tested . Leukocyte-reduced Apheresis Platelets: At least 3.0 x 1011 platelets in 90% of units tested < 5.0 x 106 WBCs 95% of units tested pH≥6.2 in at least 90% of units tested Methods o Filter: “Fourth-generation” filters remove 99.99% WBCs o Apheresis methods: most apheresis machines have built-in leukoreduction mechanisms o Less efficient methods of reducing WBC content . Washing, deglycerolizing after thawing a frozen unit, centrifugation . These methods do not meet requirement of < 5.0 x 106 WBCs per unit of RBCs/apheresis platelets. Types of leukofiltration/leukoreduction o “Pre-storage” . Done within 24 hours of collection . May use inline filters at time of collection (apheresis) or post collection o “Pre-transfusion” leukoreduction/bedside leukoreduction . Done prior to transfusion . “Bedside” leukoreduction uses gravity-based filters at time of transfusion. Least desirable given variability in practice and absence of proficiency . Alternatively performed by transfusion service prior to issuing Benefits of leukoreduction o Prevention of alloimmunization to donor HLA antigens . Anti-HLA can mediate graft rejection and immune mediated destruction of platelets o Leukoreduced products are indicated for transplant recipients or patients who are likely platelet transfusion dependent o Prevention of febrile non-hemolytic transfusion reactions (FNHTR) . -

Introduction to the Rh Blood Group.Pdf
Introduction to the Rhesus Blood Group Justin R. Rhees, M.S., MLS(ASCP)CM, SBBCM University of Utah Department of Pathology Medical Laboratory Science Program Objectives 1. Describe the major Rhesus (Rh) blood group antigens in terms of biochemical structure and inheritance. 2. Describe the characteristics of Rh antibodies. 3. Translate the five major Rh antigens, genotypes, and haplotypes from Fisher-Race to Wiener nomenclature. 4. State the purpose of Fisher-Race, Wiener, Rosenfield, and ISBT nomenclatures. Background . How did this blood group get its name? . 1937 Mrs. Seno; Bellevue hospital . Unknown antibody, unrelated to ABO . Philip Levine tested her serum against 54 ABO-compatible blood samples: only 13 were compatible. Rhesus (Rh) blood group 1930s several cases of Hemolytic of the Fetus and Newborn (HDFN) published. Hemolytic transfusion reactions (HTR) were observed in ABO- compatible transfusions. In search of more blood groups, Landsteiner and Wiener immunized rabbits with the Rhesus macaque blood of the Rhesus monkeys. Rhesus (Rh) blood group 1940 Landsteiner and Wiener reported an antibody that reacted with about 85% of human red cell samples. It was supposed that anti-Rh was the specificity causing the “intragroup” incompatibilities observed. 1941 Levine found in over 90% of erythroblastosis fetalis cases, the mother was Rh-negative and the father was Rh-positive. Rhesus macaque Rhesus (Rh) blood group Human anti-Rh and animal anti- Rh are not the same. However, “Rh” was embedded into blood group antigen terminology. The -

Cord Blood Stem Cell Transplantation
LEUKEMIA LYMPHOMA MYELOMA FACTS Cord Blood Stem Cell Transplantation No. 2 in a series providing the latest information on blood cancers Highlights • Umbilical cord blood, like bone marrow and peripheral blood, is a rich source of stem cells for transplantation. There may be advantages for certain patients to have cord blood stem cell transplants instead of transplants with marrow or peripheral blood stem cells (PBSCs). • Stem cell transplants (peripheral blood, marrow or cord blood) may use the patient’s own stem cells (called “autologous transplants”) or use donor stem cells. Donor cells may come from either a related or unrelated matched donor (called an “allogeneic transplant”). Most transplant physicians would not want to use a baby’s own cord blood (“autologous transplant”) to treat his or her leukemia. This is because donor stem cells might better fight the leukemia than the child’s own stem cells. • Cord blood for transplantation is collected from the umbilical cord and placenta after a baby is delivered. Donated cord blood that meets requirements is frozen and stored at a cord blood bank for future use. • The American Academy of Pediatrics’s (AAP) policy statement (Pediatrics; 2007;119:165-170.) addresses public and private banking options available to parents. Among several recommendations, the report encourages parents to donate to public cord blood banks and discourages parents from using private cord blood banks for personal or family cord blood storage unless they have an older child with a condition that could benefit from transplantation. • The Stem Cell Therapeutic and Research Act of 2005 put several programs in place, including creation of the National Cord Blood Inventory (NCBI) for patients in need of transplantation. -

Therapeutic Apheresis, J Clin Apheresis 2007, 22, 104-105
Apheresis: Basic Principles, Practical Considerations and Clinical Applications Joseph Schwartz, MD Anand Padmanabhan, MD PhD Director, Transfusion Medicine Assoc Med Director/Asst Prof Columbia Univ. Medical Center BloodCenter of Wisconsin New York Presbyterian Hospital Medical College of Wisconsin Review Session, ASFA Annual meeting, Scottsdale, Arizona, June 2011 Objectives (Part 1) • Mechanism of Action • Definitions • Technology (ies) • Use • Practical Considerations • Math • Clinical applications – HPC Collection Objectives (Part 2) • Clinical applications: System/ Disease Specific Indications • ASFA Fact Sheet Apheresis •Derives from Greek, “to carry away” •A technique in which whole blood is taken and separated extracorporealy, separating the portion desired from the remaining blood. •This allows the desired portion (e.g., plasma) to be removed and the reminder returned. Apheresis- Mechanism of Action •Large-bore intravenous catheter connected to a spinning centrifuge bowl •Whole blood is drawn from donor/patient into the centrifuge bowl •The more dense elements, namely the RBC, settle to the bottom with less dense elements such as WBC and platelets overlying the RBC layer and finally, plasma at the very top. Apheresis: Principles of Separation Platelets (1040) Lymphocytes Torloni MD (1050-1061) Monocytes (1065 - 1069) Granulocyte (1087 - 1092) RBC Torloni MD Torloni MD Separate blood components is based on density with removal of the desired component Graphics owned by and courtesy of Gambro BCT Principals of Apheresis WBC Plasma Torlo RBC ni MD Torloni MD RBC WBC Plasma G Cobe Spectra Apheresis- Mechanism of Action Definitions • Plasmapheresis: plasma is separated, removed (i.e. less than 15% of total plasma volume) without the use of replacement solution • Plasma exchange (TPE): plasma is separated, removed and replaced with a replacement solution such as colloid (e.g. -

Transfusion in a Rare Case of Para-Bombay Phenotype
TRANSFUSION IN A RARE CASE OF PARA-BOMBAY PHENOTYPE Charlotte Engström1, Stefan Meyer2, Young-Lan Song1, Adriana Komarek1, Alix O’Meara3, Claudia Papet3, Kathrin Neuenschwander2, Christoph Gassner2, Beat M. Frey1 1 Immunohematology, Blood Transfusion Service Zurich, Swiss Red Cross, Switzerland. 2 Molecular Diagnostics & Research, Blood Transfusion Service Zurich, Swiss Red Cross , Switzerland. 3 Hematology/Oncology, Spital Limmattal, Schlieren, Switzerland. Background Results Individuals with Bombay phenotype are characterized by the The routine anti-A, -B and -A/B failed to detect the respective absence of ABH blood group antigens both on the surface of antigens and, most notably, no H-antigen was traceable. The red blood cells (RBCs) and in secretions resulting from RBCs showed only weak agglutination with the potent anti-A/B silenced mutations in FUT1 (h/h) and FUT2 (se/se) genes, serum (Grifols). Only anti-H, but no anti-A or anti-B, was respectively. In contrast, para-Bombay phenotype retains identified in the serum. Initial ABO genotyping by sequence- some H antigen on RBCs either induced from a weakly active specific priming (PCR-SSP) resulted in AB genotype. In order (H+weak/H+weak) or completely silenced FUT1 gene (h/h). to confirm serological H-deficient phenotype a more detailed The latter is mandatory linked with an active FUT2 gene analysis was performed including sequencing of FUT1 and (Se/Se or Se/se) enabling synthesis of ABH-antigens in FUT2 which revealed an active secretor status (Se/Se) but secretions which may be adsorbed from the plasma onto homozygosity for the FUT1*01W.09 allele (c.658C>T, RBCs surface (1, 2). -

Recommendations for Collecting Red Blood Cells by Automated Apheresis Methods
Guidance for Industry Recommendations for Collecting Red Blood Cells by Automated Apheresis Methods Additional copies of this guidance document are available from: Office of Communication, Training and Manufacturers Assistance (HFM-40) 1401 Rockville Pike, Rockville, MD 20852-1448 (Tel) 1-800-835-4709 or 301-827-1800 (Internet) http://www.fda.gov/cber/guidelines.htm U.S. Department of Health and Human Services Food and Drug Administration Center for Biologics Evaluation and Research (CBER) January 2001 Technical Correction February 2001 TABLE OF CONTENTS Note: Page numbering may vary for documents distributed electronically. I. INTRODUCTION ............................................................................................................. 1 II. BACKGROUND................................................................................................................ 1 III. CHANGES FROM THE DRAFT GUIDANCE .............................................................. 2 IV. RECOMMENDED DONOR SELECTION CRITERIA FOR THE AUTOMATED RED BLOOD CELL COLLECTION PROTOCOLS ..................................................... 3 V. RECOMMENDED RED BLOOD CELL PRODUCT QUALITY CONTROL............ 5 VI. REGISTRATION AND LICENSING PROCEDURES FOR THE MANUFACTURE OF RED BLOOD CELLS COLLECTED BY AUTOMATED METHODS.................. 7 VII. ADDITIONAL REQUIREMENTS.................................................................................. 9 i GUIDANCE FOR INDUSTRY Recommendations for Collecting Red Blood Cells by Automated Apheresis Methods This -

Safe Blood and Blood Products
Safe Blood and Blood Products Module 3 Blood Group Serology Safe Blood and Blood Products Module 3 Blood Group Serology Conversion of electronic files for the website edition was supported by Cooperative Agreement Number PS001426 from the Centers for Disease Control and Prevention (CDC), Atlanta, United States of America. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of CDC. © World Health Organization, reprinted 2009 All rights reserved. Publications of the World Health Organization can be obtained from WHO Press, World Health Organization, 20 Avenue Appia, 1211 Geneva 27, Switzerland (tel.: +41 22 791 3264; fax: +41 22 791 4857; e-mail: [email protected]). Requests for permission to reproduce or translate WHO publications – whether for sale or for noncommercial distribution – should be addressed to WHO Press, at the above address (fax: +41 22 791 4806; e-mail: [email protected]). The designations employed and the presentation of the material in this publication do not imply the expression of any opinion whatsoever on the part of the World Health Organization concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Dotted lines on maps represent approximate border lines for which there may not yet be full agreement. The mention of specific companies or of certain manufacturers’ products does not imply that they are endorsed or recommended by the World Health Organization in preference to others of a similar nature that are not mentioned. Errors and omissions excepted, the names of proprietary products are distinguished by initial capital letters.