On Yield, Yield-Related Parameters and Mineral Composition of Rice (Oryza Sativa L
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Improvement of a RD6 Rice Variety for Blast Resistance and Salt Tolerance Through Marker-Assisted Backcrossing
agronomy Article Improvement of a RD6 Rice Variety for Blast Resistance and Salt Tolerance through Marker-Assisted Backcrossing Korachan Thanasilungura 1, Sukanya Kranto 2, Tidarat Monkham 1, Sompong Chankaew 1 and Jirawat Sanitchon 1,* 1 Department of Agronomy, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand; [email protected] (K.T.); [email protected] (T.M.); [email protected] (S.C.) 2 Ratchaburi Rice Research Center, Muang, Ratchaburi 70000, Thailand; [email protected] * Correspondence: [email protected]; Tel.: +66-81567-4364 Received: 24 July 2020; Accepted: 30 July 2020; Published: 1 August 2020 Abstract: RD6 is one of the most favorable glutinous rice varieties consumed throughout the north and northeast of Thailand because of its aroma and softness. However, blast disease and salt stress cause decreases in both yield quantity and quality during cultivation. Here, gene pyramiding via marker-assisted backcrossing (MAB) using combined blast resistance QTLs (qBl 1, 2, 11, and 12) and Saltol QTL was employed in solving the problem. To pursue our goal, the RD6 introgression line (RGD07005-12-165-1), containing four blast-resistant QTLs, were crossed with the Pokkali salt tolerant variety. Blast resistance evaluation was thoroughly carried out in the fields, from BC2F2:3 to BC4F4, using the upland short-row and natural field infection methods. Additionally, salt tolerance was validated in both greenhouse and field conditions. We found that the RD6 “BC4F4 132-12-61” resulting from our breeding programme successfully resisted blast disease and tolerated salt stress, while it maintained the desirable agronomic traits of the original RD6 variety. -

Agricultural and Food
REGISTERED GEOGRAPHICAL INDICATIONS INDIA- AGRICULTURAL AND FOOD S. Application Geographical Goods (As State No No. Indications per Sec 2 (f) of GIG Act 1999 ) 1 143 Guntur Sannam Chilli Agricultural Andhra Pradesh 2 121 Tirupathi Laddu Food stuff Andhra Pradesh 3 433 Bandar Laddu Food Stuff Andhra Pradesh 4 375 Arunachal Orange Agricultural Arunachal Pradesh 5 115 &118 Assam (Orthodox) Agricultural Assam 6 435 Assam Karbi Anglong Agricultural Assam Ginger 7 438 Tezpur Litchi Agricultural Assam 8 439 Joha Rice of Assam Agricultural Assam 9 558 Boka Chaul Agricultural Assam 10 609 Kaji Nemu Agricultural Assam 11 572 Chokuwa Rice of Assam Agricultural Assam 12 551 Bhagalpuri Zardalu Agricultural Bihar 13 553 Katarni Rice Agricultural Bihar 14 554 Magahi Paan Agricultural Bihar 15 552 Shahi Litchi of Bihar Agricultural Bihar 16 584 Silao Khaja Food Stuff Bihar 17 611 Jeeraphool Agricultural Chhattisgarh 18 618 Khola Chilli Agricultural Goa 19 185 Gir Kesar Mango Agricultural Gujarat 20 192 Bhalia Wheat Agricultural Gujarat 21 25 Kangra Tea Agricultural Himachal Pradesh 22 432 Himachali Kala Zeera Agricultural Himachal Pradesh 23 85 Monsooned Malabar Agricultural India Arabica Coffee (Karnataka & Kerala) 24 49 & 56 Malabar Pepper Agricultural India (Kerala, Karnataka & Tamilnadu) 25 385 Nagpur Orange Agricultural India (Maharashtra & Madhya Pradesh) 26 145 Basmati Agricultural India (Punjab / Haryana / Himachal Pradesh / Delhi / Uttarkhand / Uttar Pradesh / Jammu & Kashmir) 27 241 Banaganapalle Mangoes Agricultural India (Telangana & Andhra -

Aromatic & Medicinal Rice Varieties of Kerala.Indd
“palliyals”(single crop terrace lands ) during Rabi. pests.Sixteen species of birds have been reported from these Njavara -(Navara)-70-75 Days, Plants tall, lanky lodging paddy fi elds.The diversity of fi sh is also reported to be high in Aromatic and medicinal Panicles are short to medium in legth. Suitable for Khariff crop kundu vayals. rice varieties of Kerala in uplands and summer crop inn wetlands. Yellow and black glumed types occur. The yellow type has golden yellow lemma GANDHAKASALA ad palea white it is black in the other. Seed coats are brown in Kerala was once rich with traditional rice varieties that grow in color,endosperm non waxy and grains non-scented. Gandhakasala Rice is a long duration variety with tall plants and various agro- climatic conditions of the state. More than 2000 Chennellu -120-125 Days,Yellow and red glumed types occur. has a low grain yield of 2.0-2.7 tonnes per hectare.It is mainly traditional rice varieties including scented ad medicinal varieties The yellow type has golden yellowg grains with purple apiculus cultivated organically.Gandhakasala Rice cultivar is 150 to155cm contribute much variation to India’s bowl of rice diversity. But while the red type has bright red grains.Seed coats are brown with lesser no.of tillers (average 6 to 8 tillers) and very long many of these varieties are not cultivated any more owing to the in color,endosperm non waxy and grains non-scented.Red type panicles(27-28.4cm) with 105 to110 small, short bold ,awnless drastic reduction of paddy cultivation in the state. -

Report of Rapid Impact Assessment of Flood/ Landslides on Biodiversity Focus on Community Perspectives of the Affect on Biodiversity and Ecosystems
IMPACT OF FLOOD/ LANDSLIDES ON BIODIVERSITY COMMUNITY PERSPECTIVES AUGUST 2018 KERALA state BIODIVERSITY board 1 IMPACT OF FLOOD/LANDSLIDES ON BIODIVERSITY - COMMUnity Perspectives August 2018 Editor in Chief Dr S.C. Joshi IFS (Retd) Chairman, Kerala State Biodiversity Board, Thiruvananthapuram Editorial team Dr. V. Balakrishnan Member Secretary, Kerala State Biodiversity Board Dr. Preetha N. Mrs. Mithrambika N. B. Dr. Baiju Lal B. Dr .Pradeep S. Dr . Suresh T. Mrs. Sunitha Menon Typography : Mrs. Ajmi U.R. Design: Shinelal Published by Kerala State Biodiversity Board, Thiruvananthapuram 2 FOREWORD Kerala is the only state in India where Biodiversity Management Committees (BMC) has been constituted in all Panchayats, Municipalities and Corporation way back in 2012. The BMCs of Kerala has also been declared as Environmental watch groups by the Government of Kerala vide GO No 04/13/Envt dated 13.05.2013. In Kerala after the devastating natural disasters of August 2018 Post Disaster Needs Assessment ( PDNA) has been conducted officially by international organizations. The present report of Rapid Impact Assessment of flood/ landslides on Biodiversity focus on community perspectives of the affect on Biodiversity and Ecosystems. It is for the first time in India that such an assessment of impact of natural disasters on Biodiversity was conducted at LSG level and it is a collaborative effort of BMC and Kerala State Biodiversity Board (KSBB). More importantly each of the 187 BMCs who were involved had also outlined the major causes for such an impact as perceived by them and suggested strategies for biodiversity conservation at local level. Being a study conducted by local community all efforts has been made to incorporate practical approaches for prioritizing areas for biodiversity conservation which can be implemented at local level. -

Rice Culture in Agriculture: an Indian Perspective
The International Rice Research Institute (IRRI) was established in 1960 by the Ford and Rockefeller Foundations with the help and approval of the Government of the Philippines. Today, IRRI is one of the 15 nonprofit international research centers supported by the Consultative Group on International Agricultural Research (CGIAR - www.cgiar.org). IRRI receives support from several CGIAR members, including the World Bank, European Union, Asian Development Bank, International Fund for Agricultural Development, Rockefeller Foundation, Food and Agriculture Organization of the United Nations, and agencies of the following countries: Australia, Brazil, Canada, Denmark, France, Germany, India, Iran, Japan, Malaysia, Norway, People's Republic of China, Republic of Korea, Republic of the Philippines, Sweden, Switzerland, Thailand, United Kingdom, United States, and Vietnam. The responsibility for this publication rests with the International Rice Research Institute. © National Academy of Agricultural Sciences, New Delhi, 2007 All rights reserved. No part of this publication may be reproduced or transmitted, in any form or by any means, without permission. Any person who does any unauthorised act in relation to this publication may be liable to criminal prosecution and civil claims of damages. Mailing address: DAPO Box 7777, Metro Manila, Philippines Phone: +63 (2) 580-5600 Fax: +63 (2) 580-5699 Email: [email protected] Web: www.irri.org. Rice Knowledge Bank: www.knowledgebank.irri.org Courier address: Suite 1009, Security Bank Center 6776 Ayala Avenue, Makati City, Philippines Tel. +63 (2) 891-1236, 891-1174, 891-1258, 891-1303 Suggested Citation: Aggarwal PK, Ladha JK, Singh RK, Devakumar C, Hardy B, editors. 2007. Science, technology, and trade for peace and prosperity. -
Kuttanad Report.Pdf
Measures to Mitigate Agrarian Distress in Alappuzha and Kuttanad Wetland Ecosystem A Study Report by M. S. SWAMINATHAN RESEARCH FOUNDATION 2007 M. S. SWAMINATHAN RESEARCH FOUNDATION FOREWORD Every calamity presents opportunities for progress provided we learn appropriate lessons from the calamity and apply effective remedies to prevent its recurrence. The Alappuzha district along with Kuttanad region has been chosen by the Ministry of Agriculture, Government of India for special consideration in view of the prevailing agrarian distress. In spite of its natural wealth, the district has a high proportion of population living in poverty. The M. S. Swaminathan Research Foundation was invited by the Union Ministry of Agriculture to go into the economic and ecological problems of the Alappuzha district as well as the Kuttanad Wetland Ecosystem as a whole. The present report is the result of the study undertaken in response to the request of the Union Ministry of Agriculture. The study team was headed by Dr. S. Bala Ravi, Advisor of MSSRF with Drs. Sudha Nair, Anil Kumar and Ms. Deepa Varma as members. The Team was supported by a panel of eminent technical advisors. Recognising that the process of preparation of such reports is as important as the product, the MSSRF team held wide ranging consultations with all concerned with the economy, ecological security and livelihood security of Kuttanad wetlands. Information on the consultations held and visits made are given in the report. The report contains a malady-remedy analysis of the problems and potential solutions. The greatest challenge in dealing with multidimensional problems in our country is our inability to generate the necessary synergy and convergence among the numerous government, non-government, civil society and other agencies involved in the implementation of the programmes such as those outlined in this report. -

Registration Details of Geographical Indications
REGISTRATION DETAILS OF GEOGRAPHICAL INDICATIONS Goods S. Application Geographical Indications (As per Sec 2 (f) State No No. of GI Act 1999 ) FROM APRIL 2004 – MARCH 2005 Darjeeling Tea (word & 1 1 & 2 Agricultural West Bengal logo) 2 3 Aranmula Kannadi Handicraft Kerala 3 4 Pochampalli Ikat Handicraft Telangana FROM APRIL 2005 – MARCH 2006 4 5 Salem Fabric Handicraft Tamil Nadu 5 7 Chanderi Sarees Handicraft Madhya Pradesh 6 8 Solapur Chaddar Handicraft Maharashtra 7 9 Solapur Terry Towel Handicraft Maharashtra 8 10 Kotpad Handloom fabric Handicraft Odisha 9 11 Mysore Silk Handicraft Karnataka 10 12 Kota Doria Handicraft Rajasthan 11 13 & 18 Mysore Agarbathi Manufactured Karnataka 12 15 Kancheepuram Silk Handicraft Tamil Nadu 13 16 Bhavani Jamakkalam Handicraft Tamil Nadu 14 19 Kullu Shawl Handicraft Himachal Pradesh 15 20 Bidriware Handicraft Karnataka 16 21 Madurai Sungudi Handicraft Tamil Nadu 17 22 Orissa Ikat Handicraft Odisha 18 23 Channapatna Toys & Dolls Handicraft Karnataka 19 24 Mysore Rosewood Inlay Handicraft Karnataka 20 25 Kangra Tea Agricultural Himachal Pradesh 21 26 Coimbatore Wet Grinder Manufactured Tamil Nadu 22 28 Srikalahasthi Kalamkari Handicraft Andhra Pradesh 23 29 Mysore Sandalwood Oil Manufactured Karnataka 24 30 Mysore Sandal soap Manufactured Karnataka 25 31 Kasuti Embroidery Handicraft Karnataka Mysore Traditional 26 32 Handicraft Karnataka Paintings 27 33 Coorg Orange Agricultural Karnataka 1 FROM APRIL 2006 – MARCH 2007 28 34 Mysore Betel leaf Agricultural Karnataka 29 35 Nanjanagud Banana Agricultural -
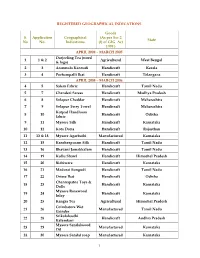
1 REGISTERED GEOGRAPHICAL INDICATIONS S. No Application No
REGISTERED GEOGRAPHICAL INDICATIONS Goods S. Application Geographical (As per Sec 2 State No No. Indications (f) of GIG Act 1999 ) APRIL 2004 – MARCH 2005 Darjeeling Tea (word 1 1 & 2 Agricultural West Bengal & logo) 2 3 Aranmula Kannadi Handicraft Kerala 3 4 Pochampalli Ikat Handicraft Telangana APRIL 2005 – MARCH 2006 4 5 Salem Fabric Handicraft Tamil Nadu 5 7 Chanderi Sarees Handicraft Madhya Pradesh 6 8 Solapur Chaddar Handicraft Maharashtra 7 9 Solapur Terry Towel Handicraft Maharashtra Kotpad Handloom 8 10 Handicraft Odisha fabric 9 11 Mysore Silk Handicraft Karnataka 10 12 Kota Doria Handicraft Rajasthan 11 13 & 18 Mysore Agarbathi Manufactured Karnataka 12 15 Kancheepuram Silk Handicraft Tamil Nadu 13 16 Bhavani Jamakkalam Handicraft Tamil Nadu 14 19 Kullu Shawl Handicraft Himachal Pradesh 15 20 Bidriware Handicraft Karnataka 16 21 Madurai Sungudi Handicraft Tamil Nadu 17 22 Orissa Ikat Handicraft Odisha Channapatna Toys & 18 23 Handicraft Karnataka Dolls Mysore Rosewood 19 24 Handicraft Karnataka Inlay 20 25 Kangra Tea Agricultural Himachal Pradesh Coimbatore Wet 21 26 Manufactured Tamil Nadu Grinder Srikalahasthi 22 28 Handicraft Andhra Pradesh Kalamkari Mysore Sandalwood 23 29 Manufactured Karnataka Oil 24 30 Mysore Sandal soap Manufactured Karnataka 1 25 31 Kasuti Embroidery Handicraft Karnataka Mysore Traditional 26 32 Handicraft Karnataka Paintings 27 33 Coorg Orange Agricultural Karnataka APRIL 2006 – MARCH 2007 28 34 Mysore Betel leaf Agricultural Karnataka 29 35 Nanjanagud Banana Agricultural Karnataka 30 37 Madhubani -

Mapping the Salt Stress-Induced Changes in the Root Mirnome in Pokkali Rice
biomolecules Article Mapping the Salt Stress-Induced Changes in the Root miRNome in Pokkali Rice Kavita Goswami 1,2, Deepti Mittal 1, Budhayash Gautam 2 , Sudhir K. Sopory 1 and Neeti Sanan-Mishra 1,* 1 Plant RNAi Biology Group, International Centre for Genetic Engineering and Biotechnology, Aruna Asaf Ali Marg, New Delhi 110067, India; [email protected] (K.G.); [email protected] (D.M.); [email protected] (S.K.S.) 2 Department of Computational Biology and Bioinformatics, Jacob School of Biotechnology and Bioengineering, Sam Higginbottom university of Agriculture, Technology and Sciences, Prayagraj (Formally Allahabad) 211007, India; [email protected] * Correspondence: [email protected]; Tel.: +91-11-2674-1358/61; Fax: +91-11-2674-2316 Received: 9 January 2020; Accepted: 6 March 2020; Published: 25 March 2020 Abstract: A plant’s response to stress conditions is governed by intricately coordinated gene expression. The microRNAs (miRs) have emerged as relatively new players in the genetic network, regulating gene expression at the transcriptional and post-transcriptional level. In this study, we performed comprehensive profiling of miRs in roots of the naturally salt-tolerant Pokkali rice variety to understand their role in regulating plant physiology in the presence of salt. For comparisons, root miR profiles of the salt-sensitive rice variety Pusa Basmati were generated. It was seen that the expression levels of 65 miRs were similar for roots of Pokkali grown in the absence of salt (PKNR) and Pusa Basmati grown in the presence of salt (PBSR). The salt-induced dis-regulations in expression profiles of miRs showed controlled changes in the roots of Pokkali (PKSR) as compared to larger variations seen in the roots of Pusa Basmati. -

Pokkali Rice Cultivation System of Kerala: an Economic Analysis
International Multidisciplinary Research Journal - ISSN 2424-7073 Agriculture & Climate Change – (July 2019): Pages 14-19 ©ICRD Publication Pokkali Rice Cultivation System of Kerala: An Economic Analysis Ranjith, P.1, K.R. Karunakaran1, S. Avudainayagam1 and A. Daniel Viswasam Samuel2 1Tamil Nadu Agricultural University University, Coimbatore, India 2Agricultural College and Research Institute Madurai, Madurai, India Abstract Pokkali rice-prawn is a unique system of rice cultivation found only in the coastal areas of Central Kerala, India where water column height of the fields increases more than 100 cm during the South-West monsoon. After the harvest of Pokkali rice- prawn stockings were introduced to these fields. It received the Geographical Indication tag in the year 2007. This research was an attempt to address production economics, efficiencies, environmental aspects etc. of this system. This study was conducted among the traditional Pokkali-prawn farmers in Ernakulum and Alappuzha districts of Kerala constituting 80 sample respondents from eight villages. Data Envelopment Analysis was used to estimate Technical (TE), Allocative (AE) and Economic Efficiencies (EE) and the Resource Use Efficiencies (RUEs) of inputs were estimated using Cobb-Douglas production function approach. The net return for the system was estimated at ₹ 281015 with Benefit-Cost ratio of 2.17. Pokkali system accounted higher efficiency measures with TE, AE and EE at 0.91, 0.97 and 0.91, respectively. The estimated RUEs of Pokkali- prawn system indicated that seeding material and labour force were over- utilized during production. Presence of NO2-N in the water samples collected from Pokkali fields indicated oxidized condition prevailed in the fields throughout the production period which reduced the methane production to minimal level. -

Paddy Through the Years Final R1.Pmd
PADDY through the years Selected writings from the Save our Rice Campaign PADDY through the years Selected writings from the Save our Rice Campaign edited by Sreedevi Lekshmikutty & Usha S PADDY Through the Years Published by Save Our Rice Campaign CREATE, 2/84, Melachatram Street, Paramakudi, Tamilnadu, India-623707 Campaign Office: Thanal, OD-3, Jawahar Nagar, Kawdiar, Thiruvananthapuram-695 003 Kerala, India. Tel. : 0471-2727150 E-mail : [email protected] Website:www.indianricecampaign.org Illustration: Dhanaraj Keezhara Layout: Corpus, Trivandrum Printing : Arsha Offset, Tvpm-1 November 2017 500 Copies West Bengal Dedicated to the memory of Nammalvar Ayya who during the Indian Workshop on Rice at Kumbalangi, 2004, said “This will make History” To Ayya, for inspiring us, motivating us and above all loving us 6 | PADDY THROUGH THE YEARS TABLE OF CONTENTS INTRODUCTION The Campaign Back Ground And The Journeys n Save Our Rice -The Campaign, The Work And Looking Ahead…..........12 n Through Karnataka In Search of Rice Seeds,Food And Farmers..............19 n Through Karnataka In Search of Rice Seeds,Food And Farmers-Part2.............24 n Save Our Rice – The Kerala Experience............31 n Journey To The Heart Of Rice Diversity..............38 n Journey Through The Delta............41 n Visit To Sagar Island, Sunderbans, West Bengal............47 Paddy Eco Systems n A Positive Water Future And Rice Cultivation In Tamil Nadu...............52 n Uncultivated Greens In A Wetland Paddy Ecosystem...............55 n Birds In Rice -

BY- ASHIRWAD SIR (Wifistudy UPSC)
2019 PIB 25.07.19 BY- ASHIRWAD SIR (wifistudy UPSC) CURRENT AFFAIRS PIB 25.07.19 BY-ASHIRWAD SIR Page 1 Cabinet approves creation of buffer stock of 40 LMT of sugar for a period of one year from 1st August 2019 to 31st July 2020 The Cabinet Committee on Economic Affairs (CCEA) chaired by Prime Minister Narendra Modi has approved the following proposals: 1. Creation of buffer stock of 40 lakh metric tonnes (LMT) of sugar for one year and to incur estimated maximum expenditure of Rs.1674 crores for this purpose. However, based on the market price and availability of sugar, this may be reviewed by the Deptt. of Food and Public Distribution any time for withdrawal / modification. 2. The reimbursement under the scheme would be met on quarterly basis to sugar mills which would be directly credited into farmers’ account on behalf of mills against cane price dues and subsequent balance, if any, would be credited to the mill’s account. Benefits: The decisions will lead to: 1. improvement in the liquidity of sugar mills; 2. reduction in sugar inventories; 3. stabilization in sugar prices by alleviating of price sentiments in domestic sugar market and thereby facilitate timely clearance of cane price dues of farmers; and 4. benefits for sugar mills in all sugarcane producing States, by way of clearing sugarcane price arrears of sugar mills Cabinet approves Determination of ‘Fair and Remunerative Price’ of sugarcane payable by sugar mills for 2019-20 sugar season The Cabinet Committee on Economic Affairs (CCEA) chaired by Prime Minister Narendra Modi has approved the proposal in respect of Determination of ‘Fair and Remunerative Price’ of sugarcane payable by sugar mills for 2019-20 sugar season.