USP42 Drives Nuclear Speckle Mrna Splicing Via Directing Dynamic Phase Separation to Promote Tumorigenesis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Computational Approach for Defining a Signature of Β-Cell Golgi Stress in Diabetes Mellitus
Page 1 of 781 Diabetes A Computational Approach for Defining a Signature of β-Cell Golgi Stress in Diabetes Mellitus Robert N. Bone1,6,7, Olufunmilola Oyebamiji2, Sayali Talware2, Sharmila Selvaraj2, Preethi Krishnan3,6, Farooq Syed1,6,7, Huanmei Wu2, Carmella Evans-Molina 1,3,4,5,6,7,8* Departments of 1Pediatrics, 3Medicine, 4Anatomy, Cell Biology & Physiology, 5Biochemistry & Molecular Biology, the 6Center for Diabetes & Metabolic Diseases, and the 7Herman B. Wells Center for Pediatric Research, Indiana University School of Medicine, Indianapolis, IN 46202; 2Department of BioHealth Informatics, Indiana University-Purdue University Indianapolis, Indianapolis, IN, 46202; 8Roudebush VA Medical Center, Indianapolis, IN 46202. *Corresponding Author(s): Carmella Evans-Molina, MD, PhD ([email protected]) Indiana University School of Medicine, 635 Barnhill Drive, MS 2031A, Indianapolis, IN 46202, Telephone: (317) 274-4145, Fax (317) 274-4107 Running Title: Golgi Stress Response in Diabetes Word Count: 4358 Number of Figures: 6 Keywords: Golgi apparatus stress, Islets, β cell, Type 1 diabetes, Type 2 diabetes 1 Diabetes Publish Ahead of Print, published online August 20, 2020 Diabetes Page 2 of 781 ABSTRACT The Golgi apparatus (GA) is an important site of insulin processing and granule maturation, but whether GA organelle dysfunction and GA stress are present in the diabetic β-cell has not been tested. We utilized an informatics-based approach to develop a transcriptional signature of β-cell GA stress using existing RNA sequencing and microarray datasets generated using human islets from donors with diabetes and islets where type 1(T1D) and type 2 diabetes (T2D) had been modeled ex vivo. To narrow our results to GA-specific genes, we applied a filter set of 1,030 genes accepted as GA associated. -

WO 2019/079361 Al 25 April 2019 (25.04.2019) W 1P O PCT
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization I International Bureau (10) International Publication Number (43) International Publication Date WO 2019/079361 Al 25 April 2019 (25.04.2019) W 1P O PCT (51) International Patent Classification: CA, CH, CL, CN, CO, CR, CU, CZ, DE, DJ, DK, DM, DO, C12Q 1/68 (2018.01) A61P 31/18 (2006.01) DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, HN, C12Q 1/70 (2006.01) HR, HU, ID, IL, IN, IR, IS, JO, JP, KE, KG, KH, KN, KP, KR, KW, KZ, LA, LC, LK, LR, LS, LU, LY, MA, MD, ME, (21) International Application Number: MG, MK, MN, MW, MX, MY, MZ, NA, NG, NI, NO, NZ, PCT/US2018/056167 OM, PA, PE, PG, PH, PL, PT, QA, RO, RS, RU, RW, SA, (22) International Filing Date: SC, SD, SE, SG, SK, SL, SM, ST, SV, SY, TH, TJ, TM, TN, 16 October 2018 (16. 10.2018) TR, TT, TZ, UA, UG, US, UZ, VC, VN, ZA, ZM, ZW. (25) Filing Language: English (84) Designated States (unless otherwise indicated, for every kind of regional protection available): ARIPO (BW, GH, (26) Publication Language: English GM, KE, LR, LS, MW, MZ, NA, RW, SD, SL, ST, SZ, TZ, (30) Priority Data: UG, ZM, ZW), Eurasian (AM, AZ, BY, KG, KZ, RU, TJ, 62/573,025 16 October 2017 (16. 10.2017) US TM), European (AL, AT, BE, BG, CH, CY, CZ, DE, DK, EE, ES, FI, FR, GB, GR, HR, HU, ΓΕ , IS, IT, LT, LU, LV, (71) Applicant: MASSACHUSETTS INSTITUTE OF MC, MK, MT, NL, NO, PL, PT, RO, RS, SE, SI, SK, SM, TECHNOLOGY [US/US]; 77 Massachusetts Avenue, TR), OAPI (BF, BJ, CF, CG, CI, CM, GA, GN, GQ, GW, Cambridge, Massachusetts 02139 (US). -

PLRG1 Antibody Cat
PLRG1 Antibody Cat. No.: 46-203 PLRG1 Antibody 46-203 (5ug/ml) staining of paraffin embedded Human Prostate. Steamed antigen retrieval with citrate buffer pH 6, AP-staining. Specifications HOST SPECIES: Goat SPECIES REACTIVITY: Human, Mouse HOMOLOGY: Expected Species Reactivity based on sequence homology: Rat, Dog, Pig, Cow IMMUNOGEN: The immunogen for this antibody is: C-PVSWKPEIIKRKRF TESTED APPLICATIONS: ELISA, IHC, WB Peptide ELISA: antibody detection limit dilution 1:16000.Western Blot:Approx 60kDa band observed in nuclear lysates of cell lines HEK293 and NIH3T3 (calculated MW of 57.2kDa according to Human NP_002660.1 and 56.9kDa according to Mouse APPLICATIONS: NP_058064.2). Recommended concentration: 0.1-0.3ug/ml. Primary incubation was 1 hour.Immunohistochemistry:Paraffin embedded Human Prostate. Recommended concentration: 5ug/ml. September 28, 2021 1 https://www.prosci-inc.com/plrg1-antibody-46-203.html This antibody is expected to recognise isoform 1 (NP_002660.1) and isoform 2 SPECIFICITY: (NP_001188493.1). POSITIVE CONTROL: 1) Cat. No. 1405 - Mouse Kidney Tissue Lysate Properties Purified from goat serum by ammonium sulphate precipitation followed by antigen PURIFICATION: affinity chromatography using the immunizing peptide. CLONALITY: Polyclonal CONJUGATE: Unconjugated PHYSICAL STATE: Liquid Supplied at 0.5 mg/ml in Tris saline, 0.02% sodium azide, pH7.3 with 0.5% bovine serum BUFFER: albumin. Aliquot and store at -20°C. Minimize freezing and thawing. CONCENTRATION: 500 ug/mL STORAGE CONDITIONS: Aliquot and store at -20˚C. -
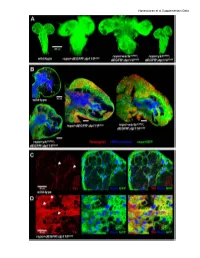
Supplementary Data Vigneswaran Et Al Supplementary Data
Vigneswaran et al Supplementary Data Vigneswaran et al Supplementary Data Figure S1: Yki is required for EGFR-PI3K-driven glial neoplasia in Drosophila (A) Optical projections of whole brain-nerve cord complexes from 3rd instar larvae approximately 130 hrs old. Dorsal view; anterior up. CD8-GFP (green) labels glial cell bodies. Compared to repo>dEGFRλ;dp110CAAX, warts knockdown (repo>wartsdsRNA; dEGFRλ;dp110CAAX) increased neoplastic brain overgrowth and yki knockdown (repo>ykidsRNA;dEGFRλ;dp110CAAX) decreased neoplastic brain overgrowth. (B) 3 µm optical projections of brain hemispheres, age-matched 3rd instar larvae. Frontal sections; anterior up; midline to left. Repo (red) labels glial cell nuclei; CD8-GFP (green) labels glial cell bodies; anti-HRP (blue) counter-stains for neurons and neuropil. (middle) repo>dEGFRλ;dp110CAAX showed increased glial cell numbers (red nuclei) compared to (upper left) wild-type. Compared to repo>dEGFRλ;dp110CAAX, (right) warts knockdown increased neoplastic glial cell numbers (red nuclei), whereas (lower left) yki knockdown reduced neoplastic glial cell numbers (red nuclei). (C, D) Low levels of Yki protein (red) was observed in wild-type central brain glia (white arrows, left panel in C) compared to high levels of cytoplasmic and nuclear Yki protein in dEGFRλ;dp110CAAX neoplastic glia (white arrows, left panel in D); Repo (blue) labels glial cell nuclei; CD8-GFP (green) labels glial cell bodies. Vigneswaran et al Supplementary Data Figure S2: YAP/TAZ expression confined to RTK-amplified tuMor cells and Maintained in patient-derived xenografts (A) On the left, immunohistochemical (IHC) staining in representative normal brain parenchyma in the cortex where YAP expression and TAZ expression was limited to vascular cells and was not detectable in normal neuronal and glial cells. -

Elucidating Biological Roles of Novel Murine Genes in Hearing Impairment in Africa
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 19 September 2019 doi:10.20944/preprints201909.0222.v1 Review Elucidating Biological Roles of Novel Murine Genes in Hearing Impairment in Africa Oluwafemi Gabriel Oluwole,1* Abdoulaye Yal 1,2, Edmond Wonkam1, Noluthando Manyisa1, Jack Morrice1, Gaston K. Mazanda1 and Ambroise Wonkam1* 1Division of Human Genetics, Department of Pathology, Faculty of Health Sciences, University of Cape Town, Observatory, Cape Town, South Africa. 2Department of Neurology, Point G Teaching Hospital, University of Sciences, Techniques and Technology, Bamako, Mali. *Correspondence to: [email protected]; [email protected] Abstract: The prevalence of congenital hearing impairment (HI) is highest in Africa. Estimates evaluated genetic causes to account for 31% of HI cases in Africa, but the identification of associated causative genes mutations have been challenging. In this study, we reviewed the potential roles, in humans, of 38 novel genes identified in a murine study. We gathered information from various genomic annotation databases and performed functional enrichment analysis using online resources i.e. genemania and g.proflier. Results revealed that 27/38 genes are express mostly in the brain, suggesting additional cognitive roles. Indeed, HERC1- R3250X had been associated with intellectual disability in a Moroccan family. A homozygous 216-bp deletion in KLC2 was found in two siblings of Egyptian descent with spastic paraplegia. Up to 27/38 murine genes have link to at least a disease, and the commonest mode of inheritance is autosomal recessive (n=8). Network analysis indicates that 20 other genes have intermediate and biological links to the novel genes, suggesting their possible roles in HI. -

Direct Interaction Between Hnrnp-M and CDC5L/PLRG1 Proteins Affects Alternative Splice Site Choice
Direct interaction between hnRNP-M and CDC5L/PLRG1 proteins affects alternative splice site choice David Llères, Marco Denegri, Marco Biggiogera, Paul Ajuh, Angus Lamond To cite this version: David Llères, Marco Denegri, Marco Biggiogera, Paul Ajuh, Angus Lamond. Direct interaction be- tween hnRNP-M and CDC5L/PLRG1 proteins affects alternative splice site choice. EMBO Reports, EMBO Press, 2010, 11 (6), pp.445 - 451. 10.1038/embor.2010.64. hal-03027049 HAL Id: hal-03027049 https://hal.archives-ouvertes.fr/hal-03027049 Submitted on 26 Nov 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. scientificscientificreport report Direct interaction between hnRNP-M and CDC5L/ PLRG1 proteins affects alternative splice site choice David Lle`res1*, Marco Denegri1*w,MarcoBiggiogera2,PaulAjuh1z & Angus I. Lamond1+ 1Wellcome Trust Centre for Gene Regulation & Expression, College of Life Sciences, University of Dundee, Dundee, UK, and 2LaboratoriodiBiologiaCellulareandCentrodiStudioperl’IstochimicadelCNR,DipartimentodiBiologiaAnimale, Universita’ di Pavia, Pavia, Italy Heterogeneous nuclear ribonucleoprotein-M (hnRNP-M) is an and affect the fate of heterogeneous nuclear RNAs by influencing their abundant nuclear protein that binds to pre-mRNA and is a structure and/or by facilitating or hindering the interaction of their component of the spliceosome complex. -

Atlas Journal
Atlas of Genetics and Cytogenetics in Oncology and Haematology Home Genes Leukemias Solid Tumours Cancer-Prone Deep Insight Portal Teaching X Y 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 NA Atlas Journal Atlas Journal versus Atlas Database: the accumulation of the issues of the Journal constitutes the body of the Database/Text-Book. TABLE OF CONTENTS Volume 12, Number 6, Nov-Dec 2008 Previous Issue / Next Issue Genes BCL8 (B-cell CLL/lymphoma 8) (15q11). Silvia Rasi, Gianluca Gaidano. Atlas Genet Cytogenet Oncol Haematol 2008; 12 (6): 781-784. [Full Text] [PDF] URL : http://atlasgeneticsoncology.org/Genes/BCL8ID781ch15q11.html CDC25A (Cell division cycle 25A) (3p21). Dipankar Ray, Hiroaki Kiyokawa. Atlas Genet Cytogenet Oncol Haematol 2008; 12 (6): 785-791. [Full Text] [PDF] URL : http://atlasgeneticsoncology.org/Genes/CDC25AID40004ch3p21.html CDC73 (cell division cycle 73, Paf1/RNA polymerase II complex component, homolog (S. cerevisiae)) (1q31.2). Leslie Farber, Bin Tean Teh. Atlas Genet Cytogenet Oncol Haematol 2008; 12 (6): 792-797. [Full Text] [PDF] URL : http://atlasgeneticsoncology.org/Genes/CDC73D181ch1q31.html EIF3C (eukaryotic translation initiation factor 3, subunit C) (16p11.2). Daniel R Scoles. Atlas Genet Cytogenet Oncol Haematol 2008; 12 (6): 798-802. [Full Text] [PDF] URL : http://atlasgeneticsoncology.org/Genes/EIF3CID44187ch16p11.html ELAC2 (elaC homolog 2 (E. coli)) (17p11.2). Yang Chen, Sean Tavtigian, Donna Shattuck. Atlas Genet Cytogenet Oncol Haematol 2008; 12 (6): 803-806. [Full Text] [PDF] URL : http://atlasgeneticsoncology.org/Genes/ELAC2ID40437ch17p11.html FOXM1 (forkhead box M1) (12p13). Jamila Laoukili, Monica Alvarez Fernandez, René H Medema. -

RUNX1-3′USP42 Chimeric Gene in Acute Myeloid Leukemia Can Occur
Zagaria et al. Molecular Cytogenetics 2014, 7:66 http://www.molecularcytogenetics.org/content/7/1/66 CASE REPORT Open Access 5′RUNX1-3′USP42 chimeric gene in acute myeloid leukemia can occur through an insertion mechanism rather than translocation and may be mediated by genomic segmental duplications Antonella Zagaria, Luisa Anelli, Nicoletta Coccaro, Giuseppina Tota, Paola Casieri, Angelo Cellamare, Angela Minervini, Crescenzio Francesco Minervini, Claudia Brunetti, Cosimo Cumbo, Giorgina Specchia and Francesco Albano* Abstract Background: The runt-related transcription factor 1 (RUNX1) gene is a transcription factor that acts as a master regulator of hematopoiesis and represents one of the most frequent targets of chromosomal rearrangements in human leukemias. The t(7;21)(p22;q22) rearrangement generating a 5′RUNX1-3′USP42 fusion transcript has been reported in two cases of pediatric acute myeloid leukemia (AML) and further in eight adult cases of myeloid neoplasms. We describe the first case of adult AML with a 5′RUNX1-3′USP42 fusion gene generated by an insertion event instead of chromosomal translocation. Methods: Conventional and molecular cytogenetic analyses allowed the precise characterization of the chromosomal rearrangement and breakpoints identification. Gene expression analysis was performed by quantitative real-time PCR experiments, whereas bioinformatic studies were carried out for revealing structural genomic characteristics of breakpoint regions. Results: We identified an adult AML case bearing a ins(21;7)(q22;p15p22) generating a 5′RUNX1-3′USP42 fusion gene on der(21) chromosome and causing USP42 gene over-expression. Bioinformatic analysis of the genomic regions involved in ins(21;7)/t(7;21) showed the presence of interchromosomal segmental duplications (SDs) next to the USP42 and RUNX1 genes, that may underlie a non-allelic homologous recombination between chromosome 7 and 21 in AML. -

Mrna Editing, Processing and Quality Control in Caenorhabditis Elegans
| WORMBOOK mRNA Editing, Processing and Quality Control in Caenorhabditis elegans Joshua A. Arribere,*,1 Hidehito Kuroyanagi,†,1 and Heather A. Hundley‡,1 *Department of MCD Biology, UC Santa Cruz, California 95064, †Laboratory of Gene Expression, Medical Research Institute, Tokyo Medical and Dental University, Tokyo 113-8510, Japan, and ‡Medical Sciences Program, Indiana University School of Medicine-Bloomington, Indiana 47405 ABSTRACT While DNA serves as the blueprint of life, the distinct functions of each cell are determined by the dynamic expression of genes from the static genome. The amount and specific sequences of RNAs expressed in a given cell involves a number of regulated processes including RNA synthesis (transcription), processing, splicing, modification, polyadenylation, stability, translation, and degradation. As errors during mRNA production can create gene products that are deleterious to the organism, quality control mechanisms exist to survey and remove errors in mRNA expression and processing. Here, we will provide an overview of mRNA processing and quality control mechanisms that occur in Caenorhabditis elegans, with a focus on those that occur on protein-coding genes after transcription initiation. In addition, we will describe the genetic and technical approaches that have allowed studies in C. elegans to reveal important mechanistic insight into these processes. KEYWORDS Caenorhabditis elegans; splicing; RNA editing; RNA modification; polyadenylation; quality control; WormBook TABLE OF CONTENTS Abstract 531 RNA Editing and Modification 533 Adenosine-to-inosine RNA editing 533 The C. elegans A-to-I editing machinery 534 RNA editing in space and time 535 ADARs regulate the levels and fates of endogenous dsRNA 537 Are other modifications present in C. -

Structural Insights Into Nuclear Pre-Mrna Splicing in Higher Eukaryotes
Downloaded from http://cshperspectives.cshlp.org/ on September 28, 2021 - Published by Cold Spring Harbor Laboratory Press Structural Insights into Nuclear pre-mRNA Splicing in Higher Eukaryotes Berthold Kastner,1 Cindy L. Will,1 Holger Stark,2 and Reinhard Lührmann1 1Department of Cellular Biochemistry, Max Planck Institute for Biophysical Chemistry, D-37077 Göttingen, Germany 2Department of Structural Dynamics, Max Planck Institute for Biophysical Chemistry, D-37077 Göttingen, Germany Correspondence: [email protected] SUMMARY The spliceosome is a highly complex, dynamic ribonucleoprotein molecular machine that undergoes numerous structural and compositional rearrangements that lead to the formation of its active site. Recent advances in cyroelectron microscopy (cryo-EM) have provided a plethora of near-atomic structural information about the inner workings of the spliceosome. Aided by previous biochemical, structural, and functional studies, cryo-EM has confirmed or provided a structural basis for most of the prevailing models of spliceosome function, but at the same time allowed novel insights into splicing catalysis and the intriguing dynamics of the spliceosome. The mechanism of pre-mRNA splicing is highly conserved between humans and yeast, but the compositional dynamics and ribonucleoprotein (RNP) remodeling of the human spliceosome are more complex. Here, we summarize recent advances in our understanding of the molec- ular architecture of the human spliceosome, highlighting differences between the human and yeast -

Downregulation of Carnitine Acyl-Carnitine Translocase by Mirnas
Page 1 of 288 Diabetes 1 Downregulation of Carnitine acyl-carnitine translocase by miRNAs 132 and 212 amplifies glucose-stimulated insulin secretion Mufaddal S. Soni1, Mary E. Rabaglia1, Sushant Bhatnagar1, Jin Shang2, Olga Ilkayeva3, Randall Mynatt4, Yun-Ping Zhou2, Eric E. Schadt6, Nancy A.Thornberry2, Deborah M. Muoio5, Mark P. Keller1 and Alan D. Attie1 From the 1Department of Biochemistry, University of Wisconsin, Madison, Wisconsin; 2Department of Metabolic Disorders-Diabetes, Merck Research Laboratories, Rahway, New Jersey; 3Sarah W. Stedman Nutrition and Metabolism Center, Duke Institute of Molecular Physiology, 5Departments of Medicine and Pharmacology and Cancer Biology, Durham, North Carolina. 4Pennington Biomedical Research Center, Louisiana State University system, Baton Rouge, Louisiana; 6Institute for Genomics and Multiscale Biology, Mount Sinai School of Medicine, New York, New York. Corresponding author Alan D. Attie, 543A Biochemistry Addition, 433 Babcock Drive, Department of Biochemistry, University of Wisconsin-Madison, Madison, Wisconsin, (608) 262-1372 (Ph), (608) 263-9608 (fax), [email protected]. Running Title: Fatty acyl-carnitines enhance insulin secretion Abstract word count: 163 Main text Word count: 3960 Number of tables: 0 Number of figures: 5 Diabetes Publish Ahead of Print, published online June 26, 2014 Diabetes Page 2 of 288 2 ABSTRACT We previously demonstrated that micro-RNAs 132 and 212 are differentially upregulated in response to obesity in two mouse strains that differ in their susceptibility to obesity-induced diabetes. Here we show the overexpression of micro-RNAs 132 and 212 enhances insulin secretion (IS) in response to glucose and other secretagogues including non-fuel stimuli. We determined that carnitine acyl-carnitine translocase (CACT, Slc25a20) is a direct target of these miRNAs. -

Alternatively Spliced Genes
1 Alternatively Spliced Genes Jane Y. Wu1,2,LiyaYuan1 and Necat Havlioglu1 1Washington University School of Medicine, St. Louis, MO, USA 2John F. Kennedy Center for Research on Human Development, Vanderbilt University Medical Center, Nashville, TN, USA 1 Pre-mRNA Splicing and Splicing Machinery 3 1.1 Splicing Machinery: Spliceosome 3 1.2 Splicing Signals 4 1.3 Spliceosomal UsnRNP Biogenesis 12 1.4 Spliceosome Assembly 13 1.5 Biochemical Mechanisms of pre-mRNA Splicing 17 2 Alternative pre-mRNA Splicing 17 2.1 Alternative Splicing and its Role in Regulating Gene Activities and Generating Genetic Diversity 17 2.1.1 Different Patterns of Alternative Splicing 17 2.1.2 Alternative Splicing and Genetic Diversity 18 2.2 Mechanisms Underlying Alternative Splicing Regulation 19 2.2.1 Splicing Signals and Splicing Regulatory Elements 20 2.2.2 Trans-acting Splicing Regulators 23 2.3 Tissue-specific and Developmentally Regulated Alternative Splicing 26 2.4 Regulation of Alternative Splicing in Response to Extracellular Stimuli 27 3 Pre-mRNA Splicing and Human Diseases 28 3.1 Splicing Defects in Human Diseases 28 3.2 Molecular Mechanisms Underlying Splicing Defects Associated with Disease 33 4 Perspectives on Diagnosis and Treatment of Diseases Caused by pre-mRNA Splicing Defects 36 4.1 Diagnosis of Human Diseases Caused by Splicing Defects 36 2 Alternatively Spliced Genes 4.2 Potential Therapeutic Approaches 37 4.2.1 Oligonucleotide-based Approaches: Antisense, RNAi, and Chimeric Molecules 37 4.2.2 Ribozymes 37 4.2.3 SMaRT 38 4.2.4 Chemical Compounds 38 5 Concluding Remarks 38 Acknowledgment 39 Bibliography 39 Books and Reviews 39 Keywords Pre-mRNA Nascent transcripts that are precursors of mature messenger RNAs.