Running Head
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

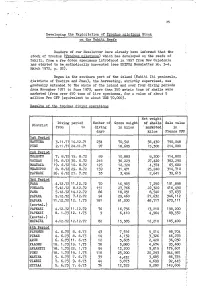
Répartition De La Population En Polynésie Française En 2017
Répartition de la population en Polynésie française en 2017 PIRAE ARUE Paopao Teavaro Hatiheu PAPEETE Papetoai A r c h MAHINA i p e l d FAA'A HITIAA O TE RA e s NUKU HIVA M a UA HUKA r q PUNAAUIA u HIVA OA i TAIARAPU-EST UA POU s Taiohae Taipivai e PAEA TA HUATA s NUKU HIVA Haapiti Afareaitu FATU HIVA Atuona PAPARA TEVA I UTA MOO REA TAIARAPU-OUEST A r c h i p e l d Puamau TAHITI e s T MANIHI u a HIVA OA Hipu RA NGIROA m Iripau TA KAROA PUKA P UKA o NA PUKA Hakahau Faaaha t u Tapuamu d e l a S o c i é MAKEMO FANGATA U - p e l t é h i BORA BORA G c a Haamene r MAUPITI Ruutia A TA HA A ARUTUA m HUAHINE FAKARAVA b TATAKOTO i Niua Vaitoare RAIATEA e TAHITI r TAHAA ANAA RE AO Hakamaii MOORE A - HIK UE RU Fare Maeva MAIAO UA POU Faie HA O NUKUTAVAKE Fitii Apataki Tefarerii Maroe TUREIA Haapu Parea RIMATARA RURUTU A r c h Arutua HUAHINE i p e TUBUAI l d e s GAMBIE R Faanui Anau RA IVAVAE A u s Kaukura t r Nombre a l AR UTUA d'individus e s Taahuaia Moerai Mataura Nunue 20 000 Mataiva RA PA BOR A B OR A 10 000 Avera Tikehau 7 000 Rangiroa Hauti 3 500 Mahu Makatea 1 000 RURUT U TUBUAI RANGIROA ´ 0 110 Km So u r c e : Re c en se m en t d e la p o p u la ti o n 2 0 1 7 - IS P F -I N SE E Répartition de la population aux Îles Du Vent en 2017 TAHITI MAHINA Paopao Papetoai ARUE PAPEETE PIRAE HITIAA O TE RA FAAA Teavaro Tiarei Mahaena Haapiti PUNAAUIA Afareaitu Hitiaa Papenoo MOOREA 0 2 Km Faaone PAEA Papeari TAIARAPU-EST Mataiea Afaahiti Pueu Toahotu Nombre PAPARA d'individus TEVA I UTA Tautira 20 000 Vairao 15 000 13 000 Teahupoo 10 000 TAIARAPU-OUEST -
Developing the Exploitation of <I>Trochus Niloticus</I> Stock on the Tahiti Reefs
35 Developing the Exploitation of Trochu3 niloticus Stock onithe_Ta'hiti Reefs Readers of our Newsletter have already been informed that the stock of trochus (Trochug niloticus) which has developed on the reefs of Tahiti, from a few dozen specimens introduced in 1957 from Hew Caledonia has started to be methodically harvested (see SPIPDA Newsletter ITo. 3-4, March 1972, p. 32). Begun in the southern part of the island (Tahiti Iti peninsula, districts of Tautira and Pueu), the harvesting, strictly supervised, was gradually extended to the whole of the island and over four diving periods fjrom November 1971 to June 1973? more than 350 metric tons of shells were marketed (from over 450 tons of live specimens, for a value of about 5 million Frs CTP (equivalent to about US$ 70,000). Results of .the trochus. diving operations Net weight Diving period Number of Gross weight of shells Sale value District from to diving in kilos ' marketed in days kilos Francs CPP 1st Period TAUTIRA 3-11.71 I4.l2.7i 234 70,541 56,430 790,048 .. PUEU 2.11 .71 24.11.71 97 18,605 ... 15,300 214,000 2nd Period TOAHOTU 7. 8.72 15. 8,72 89 10r883 8,200 114,800 : VAIRAO 15. 8.72 30. 8.72 216 36J229 27,420 362,290 , MAATAIA. 19. 6.72 10. 8o72 125 12,724 4,374 65,600 TEAHUPOO 8. 8.72 29. 8.72 159 31,471 25,240' 314,710 PAPEARI 26. 6.72 27. 7.72 35 3,456 2,641 39,615 ; 3rd Period FAAA 4.12,72 11.12.72 70 16,983 7,250 101,898 PIMAAUA 5.12.72 8,12.72 151 27,766 .22,320 416,490 PAEA 5.12.72 14*12.72 86 18,051 8, 540 97,633 PAPARA 5.12.72 7.12.72 94 29,460 21,632 346,112 PAPARA 11.12.72 12. -

Bioerosion of Experimental Substrates on High Islands and on Atoll Lagoons (French Polynesia) After Two Years of Exposure
MARINE ECOLOGY PROGRESS SERIES Vol. 166: 119-130,1998 Published May 28 Mar Ecol Prog Ser -- l Bioerosion of experimental substrates on high islands and on atoll lagoons (French Polynesia) after two years of exposure 'Centre d'oceanologie de Marseille, UMR CNRS 6540, Universite de la Mediterranee, Station Marine d'Endoume, rue de la Batterie des Lions, F-13007 Marseille, France 2Centre de Sedimentologie et Paleontologie. UPRESA CNRS 6019. Universite de Provence, Aix-Marseille I, case 67, F-13331 Marseille cedex 03. France 3The Australian Museum, 6-8 College Street, 2000 Sydney, New South Wales, Australia "epartment of Biology, Boston University, Boston, Massachusetts 02215, USA ABSTRACT: Rates of bioerosion by grazing and boring were studied in lagoons of 2 high islands (3 sites) and 2 atolls (2 sites each) In French Polynesia using experimental carbonate substrates (blocks of Porites lutea skeleton). The substrate loss versus accretion was measured after 6 and 24 mo of exposure. The results show significant differences between pristine environments on atolls and envi- ronments on high islands sub!ected to different levels of eutrophication and pollution due to human activities. Whereas experimental substrates on the atolls maintain a balance between accretion and erosion or exhibit net gains from accretion (positive budget), only 1 site on a high island exhibits sig- nificant loss of substrate by net erosion (negative budget). The erosional patterns set within the first 6 rno of exposure were largely maintained throughout the entire duration of the expeiiment. The inten- sity of bioerosion by grazing increases dramatically when reefs are exposed to pollution from harbour waters; this is shown at one of the Tahiti sites, where the highest average bioerosional loss, up to 25 kg m-2 yr-' (6.9 kg m-' yr-I on a single isolated block), of carbonate substrate was recorded. -

Liste Des Labellisés CLCE Au 30 Novembre 2017 Arue Faaa
Liste des Labellisés CLCE au 30 novembre 2017 TAHITI Arue ➢ DOMOTEC Contact : CHOUGUES Gilles 87 78 15 35 40 45 34 07 [email protected] Courrier : BP 14717 – 98701 ARUE Faaa 1 ➢ 987 ELEC Contact : WATANABE Otis 87 78 43 22 : 40 82 30 71 [email protected] Courrier : BP 61609 – 98702FAA’A ➢ ARESO Contact : MOUSSET Laurent 87 70 73 42 [email protected] Courrier : BP 61781 – 98704 FAA’A Centre ➢ ELEC 220 Contact : HERLEMME Mickael 87 74 73 09 [email protected] Courrier : BP 3341 – 98717 Punaauia Association Centre Label et Contrôle Electrique Téléphone : 40 47 27 72 Liste des Labellisés CLCE au 30 novembre 2017 ➢ TEARATAI UIRA Contact : KONG YEK FHAN Christian 87 72 59 21 [email protected] Courrier : BP 6129 – 98704 FAA’A ➢ TECHNO FROID Contact : LANVIN Jérôme 40 80 04 05 40 80 04 09 [email protected] Courrier : BP 50354 – 98716 PIRAE Contact : TEHURITAUA Marc ➢ TEHURITAUA & FILS 2 87 77 36 21 [email protected] Courrier : BP 6276 – 98702 FAA’A Mahina ➢ A.E.S ELECTRICITE Contact : MICHON Nicolas 87 34 34 87 [email protected] Courrier : BP 90124 Motu Uta – 98715 Papeete ➢ POLY RESEAUX CONCEPT Contact : OPUTU John 87 34 63 24 [email protected] Association Centre Label et Contrôle Electrique Téléphone : 40 47 27 72 Liste des Labellisés CLCE au 30 novembre 2017 Paea ➢ ELECTRICITE ET RESEAUX DE Contact : LALANDEC Patrick TAHITI (ERDT) 87 77 09 83 / 87 78 75 31 40 50 79 41 [email protected] Courrier : BP 50131 – 98716 PIRAE ➢ WES ELEC Contact : BUTSCHER Wesley 89 72 78 31 / 87 75 08 15 40 81 34 66 [email protected] / [email protected] -

Partial Flora of the Society Islands: Ericaceae to Apocynaceae
SMITHSONIAN CONTRIBUTIONS TO BOTANY NUMBER 17 Partial Flora of the Society Islands: Ericaceae to Apocynaceae Martin Lawrence Grant, F. Raymond Fosberg, and Howard M. Smith SMITHSONIAN INSTITUTION PRESS City of Washington 1974 ABSTRACT Grant, Martin Lawrence, F. Raymond Fosberg, and Howard M. Smith. Partial Flora of the Society Islands: Ericaceae to Apocynaceae. Smithsonian Contri- butions to Botany, number 17, 85 pages, 1974.-Results of a botanical inves- tigation of the Society Islands carried out by Grant in 1930 and 1931, and subsequent work on the material collected and other collections in the U.S. herbaria and other published works are reported herein. This paper is a partial descriptive flora of the Society group with a history of the botanical exploration and investigation of the area. OFFICIALPUBLICATION DATE is handstamped in a limited number of initial copies and is recorded in the Institution’s annual report, Srnithsonian Year. SI PRESS NUMBER 5056. SERIES COVER DESIGN: Leaf clearing from the katsura tree Cercidiphyllurn juponicum Siebold and Zuccarini. Library of Congress Cataloging in Publication Data Grant, Martin Lawrence, 1907-1968. Partial flora of the Society Islands: Ericaceae to Apocynaceae. (Smithsonian contributions to botany, no. 17) Supt. of Docs. no.: SI 1.29:17. 1. Botany-Society Islands. I. Fosberg, Francis Raymond, 1908- , joint author. 11. Smith, Howard Malcolm, 1939- , joint author. 111. Title. IV. Series: Smithsonian Institution. Smith- sonian contributions to botany, no. 17. QK1.2747 no. 17 581’.08s [581.9’96’21] 73-22464 For sale by the Superintendent of Documents, US. Government Printing Office Washington, D.C. 20402 Price $1.75 (paper cover) The senior author, after spending almost a year during 1930 and 1931 in the Society Islands, collecting herbarium material and ecological data, worked inten- sively on a comprehensive flora of this archipelago for the next five years. -

The Pacific Islands
American Universities Field Staff THIS FIELDSTAFF REPORT is one of a continuing series The on international affairs and major global issues of our time. Fieldstaff Reports have for twenty-five years American reached a group of readers--both academic and non- academic--who find them a useful source of firsthand Universities observation of political, economic, and social trends in foreign countries. Reports in the series are prepared by Field Staff writers who are full-time Associates of the American Universities Field Staff and occasionally by persons on P.O. Box 150, Hanover, NH 03755 leave from the organizations and universities that are the Field Staff's sponsors. Associates of the Field Staff are chosen for their ability to cut across the boundaries of the academic disciplines in order to study societies in their totality, and for their skill in collecting, reporting, and evaluating data. They combine long residence abroad with scholarly studies relating to their geographic areas of interest. Each Field Staff Associate returns to the United States periodically to lecture on the campuses of the consortium's member institutions. The American Universities Field Staff, Inc., founded in 1951 as a nonprofit organization of American educational institutions, engages in various international activities both at home and in foreign areas. These activities have a wide range and include writing on social and political change in the modern world, the making of documentary films (Faces of Change), and the organizing of seminars and teaching of students at the Center for Mediterranean Studies in Rome and the Center for Asian and Pacific Studies in Singapore. -

Power Switching and Renewal in French Polynesian Politics 1
Power switching and renewal in French Polynesian politics 1 Power switching and renewal in French Polynesian politics the importance of 2004 Bruno Saura translated by Bess Flores Abstract The year 2004 saw great upheavals in the political situation in Tahiti, and may prove to be a very important year in the history of French Polynesia as a whole. The May elections for the Territorial Assembly, and thus for the President of the territory, resulted for the first time in the victory of Oscar Temaru. A long-time challenger of the outgoing President Gaston Flosse, Temaru had suspended his programme of immediate independence in favour of an ideological breathing space of several years, with two aims: to break Gaston Flosse’s autocratic grip on power, and to set up real and sustainable development processes. Oscar Temaru became President at the head of an unstable majority in May 2004 but lost power to Gaston Flosse in October 2004. He was re-elected, again with a fragile majority, in the February 2005 elections. This article, based on the facts of 2004 and early 2005, reveals the metropolitan French government’s lack of neutrality in the electoral process (notably by adopting a system intended to favour Gaston Flosse). It examines the relationship between politics and religion in French Polynesia; highlights cultural factors in French Polynesian politics, notes the emergence of young potential leaders, and thereby explains the reasons for the political rise and fall and rise again of Oscar Temaru. Keywords elections; French Polynesia; politics and religion; Tahiti. The Journal of Pacific Studies, Volume 28, no.1, 2005, 1–22 © by JPacS Editorial Board (SSED,USP) 1 2 The Journal of Pacific Studies Vol.28 no.1, 2005 Résumé L’année politique 2004 à Tahiti a connu de grands bouleversements. -

Classical Biological Control of the Glassy-Winged Sharpshooter
Biol Invasions (2008) 10:135–148 DOI 10.1007/s10530-007-9116-y ORIGINAL PAPER Engineering an invasion: classical biological control of the glassy-winged sharpshooter, Homalodisca vitripennis, by the egg parasitoid Gonatocerus ashmeadi in Tahiti and Moorea, French Polynesia Julie Grandgirard Æ Mark S. Hoddle Æ Jerome N. Petit Æ George K. Roderick Æ Neil Davies Received: 13 October 2006 / Accepted: 19 April 2007 / Published online: 25 May 2007 Ó Springer Science+Business Media B.V. 2007 Abstract The glassy-winged sharpshooter, Homa- surveys during the first year of their interaction in lodisca vitripennis Germar (=H. coagulata Say) French Polynesia (until mid-May 2006). The impact (Hemiptera: Cicadellidae), invaded Tahiti in 1999 of G. ashmeadi on H. vitripennis was extremely rapid and spread rapidly to the main island groups of and high. Parasitism of H. vitripennis egg masses by French Polynesia becoming an important pest. It G. ashmeadi has averaged 80–100% in Tahiti since threatened agriculture, native biodiversity, and the introduction of the parasitoid, and populations of created serious social and recreational problems. H. vitripennis nymphs and adults have decreased by Further, massive uncontrolled populations on Tahiti more than 90% since December 2005. Populations presented an elevated invasion threat to other South of H. vitripennis have been successfully maintained Pacific nations. In 2004, a classical biological control at this low level for more than 1 year. The same program against H. vitripennis was initiated in French results were obtained in nearby Moorea where the Polynesia using the highly host-specific egg parasit- parasitoid was probably spread by the unregulated oid Gonatocerus ashmeadi Girault (Hymenoptera: transport of plants infested with parasitized H. -

29-Sep-2015 2 Passengers in Economy Class on Air France Flight
Reservation Status : Confirmed Reservation Date : 28-Apr-2015 Departure Date : 29-Sep-2015 Itinerary Date : 04-Aug-2015 29-Sep-2015 2 Passengers in Economy Class on Air France Flight 76 Tuesday Departing Los Angeles, California airport at 11:55P Passenger 1 PNR 69WWN6 Seat 30C Passenger 2 PNR 69WWN6 Airport check-in for seat assignment Passenger 1 Ticket# 0577646744097 E-Ticket Passenger 2 Ticket# 0577646744099 E-Ticket reservations are made. 30-Sep-2015 2 Passengers in Economy Class on Air France Flight 76 Wednesday Arriving Papeete, Tahiti airport at 5:10A Passenger 1 PNR 69WWN6 Seat 30C Passenger 2 PNR 69WWN6 Airport check-in for seat assignment 2 Transfers From: Papeete, Tahiti Airport To: Papeete, Tahiti Pier Transfer A lei greeting is included for guests arriving on international flights. Surfboard transport is not included. Arrangements must be made in advance for an additional fee. Please inquire for further information. 2 Passengers in Coach Class on Terevau Catamaran Ship 18 Departing Papeete, Tahiti pier at 8:10A Arriving Temae, Moorea pier at 8:40A Passengers 1-2 Pleasant Holidays Res #1956687 Page ..... 2 30-Sep-2015 2 Transfers Wednesday From: Temae, Moorea Pier To: Hilton Moorea Lagoon Resort Transfer Hilton Moorea Lagoon Resort & Spa 1 Lagoon Bungalow Room Early Bird Offer for 4 Nights for 2 Guests Guests 1-2 PO Box 1005 Papetoai, MO PYF 98728 Room Number 1 Confirmation 3191569693 Breakfast Plan CHECK IN: 2:00PM CHECK OUT: 11:00AM 1 One Black Pearl Pendant in Temae, Moorea 04-Oct-2015 2 Transfers Sunday From: Hilton Moorea Lagoon Resort To: Temae, Moorea Airport Transfer 2 Passengers on Air Tahiti Flight 456(1 stop) Departing Temae, Moorea airport at 9:15A Arriving Bora Bora, Motu Mute airport at 10:30A Passengers 1-2 PNR P0YJM Airport check-in for seat assignment 2 Transfers From: Bora Bora, Motu Mute Airport To: IC Bora Bora Le Moana Resort Transfer Pleasant Holidays Res #1956687 Page .... -

A Survey of the Economy of French Polynesia 1960 to 1990
Islands/Australia Working Paper No. 91/4 A survey of the economy of French Polynesia 1960 to 1990 Gilles Blanchet A a S 001 f Pacifi S dIe ISSN 0816-5165 ISBN 0 7315 0937 4 Islands!Australia Working Paper No. 91!4 A survey of the economy of French Polynesia 1960 to 1990 Gilles Blanchet This series is intended to provide prompt preliminary distribution of new work on development studies to interested scholars. It is complementary to the Centre's Monograph series, and intended primarily for shorter pieces and for work that is not yet in its final form. The views expressed in this Working Paper are those of the author and should not be attributed to the National Centre for Development Studies. © National Centre for Development Studies 1991 This work is copyright. Apart from those uses which may be permitted under the Copyright Act 1968 as amended, no part may he reproduced by any process without written permission. Inquiries should be made to the publisher. Gilles Blanchet is an economic and sociological researcher at ORSTROM, now known as the Institut Français de Recherche Scientifique pour le Développement en Coopération. He has worked for several years in French Polynesia where he was involved in small-seale fishery studies. He was a Visiting Fellow at the National Centre for Development Studies, Austmlian National University, until July 1991. Key to symbols used in tables n.a Not applicable Not available Zero Insignificant Printed in Australia by Soepae Prinlery National Centre for Development Studies Research School ofPacifie Studies The Australian National University GPO Box 4 Canberra ACT 2601 Australia Telephone (06) 2494705 EaJl;(06) 2572886 Abstract This paper is a shortened and updated version of a previous report on French Polynesia's economy (Blanchet 1985), and is a descriptive overview rather than a comprehensive survey. -

Entrepreneurs De Taxi
ENTREPRENEURS DE TAXI ÎLES DU VENT TAHITI ADRESSE ENTREPRENEUR ADRESSE GEOGRAPHIQUE TEL FAX EMAIL POSTALE Paea - PK 28 c/mont Maraa - AIRIMA Henry 98711 Paea 87 77 52 39 - - Servitude Stuart AITAMAI Marie, Faa'a - PK 5,500 c/mont - Face BP 61156 87 77 82 82 - - Sidonie, Tina aéroport 98702 Faa'a BP 12618 AKA Justin PK 39,500 c/mont 87 20 80 88 - - 98712 Papara Punaauia - PK 12,5 c/mont BP 20386 40 43 82 21 ATAE Fred - - Qtier Deligny 3 98713 Papeete 87 77 87 12 AUTAI Sonny, BP 61903 Faa'a - Pamatai - Qtier Hugonot 87 29 88 40 - - Teva 98702 Faa'a BP 381249 40 58 38 42 BENNETT Marcel Punaauia - PK 16,8 c/mer - - 98718 Punaauia 87 77 37 19 BP 5705 BENNETT Paulette Pirae - Hamuta, plateau n° 45 87 77 17 77 - - 98716 Pirae BROTHERSON BP 120055 40 82 83 76 Papara - PK 32 c/mer - [email protected] Steve, Tuarii 98712 Papara 87 74 16 67 BRUTO Tihoti, BP 6284 Faa'a - Puurai Lot N° 84 87 79 37 62 - - Dick 98702 Faa'a CAMPS épse TERIIPAIA Punaauia - Lotissement Miri - BP 50090 40 85 36 03 - [email protected] Ghislaine, Immeuble Eeva - Appart. A 98716 Pirae 87 72 22 94 Jacqueline BP 14564 CARLSON Karl Arue - PK 3 c/mont face Carrefour 87 78 73 19 - 98701 Arue - Papeete - Mission - Lotissement BP 62323 40 43 47 68 CHISAKA Yoshiaki - [email protected] Tepapa n° 1 98703 Faa'a 87 72 80 30 CHONG Amani, BP 2506 Faa'a - Puurai - Qtier Bonnefin 87 79 75 28 - - Teurukura 98713 Papeete COLOMBEL Faa'a - PK 4 c/mont - Qtier BP 9973 40 81 98 83 - - Robert Colombel 98715 Papeete 87 77 98 19 DEXTER Ernest, BP 130308 40 45 54 32 Faa'a - Résidence Manutea -

Society Islands Insects
SOCIETY ISLANDS INSECTS BERNICE P. BISHOP MUSEUM BULLETIN 113 PACIFIC ENTOMOLOGICAL SURVEY PUBLICATION 6 HONOLULU, HAWAII PUBLISHED BY THE MUSEUM 1935 SOCIETY ISLANDS INSECTS CONTENTS ARTIeI.E PAGE 1. Termites from the Society Islands, by S. F. Light (1 figure, 1 plate).................... 3 2. Mallophaga from Tahiti, by G. F. Ferris (4 figures) --...................... 7 3. Ants from the Society Islands, by William Morton Wheeler.. h __ .__ 13 4. Odonata from Tahiti, by James G. Needham (1 figure) 00 ·........ 21 5. Chermidae from the Society Islands, by F. D. Klyver (1 figure) 25 6. Thysanoptera from the Society Islands, by Dudley Moulton (2 figures)................ 29 7. Tahitian Simuliidae, by F. W. Edwards (2 figures) .. __ __ .__ .__ 35 8. Cercopidae of the Society Islands, by W. E. China (6 figures) 00 ..00........... 39 9. Rhyncogonus submetallicus, new species, from Tahiti, by Edwin C. Van Dyke (1 figure) .. 00 ·•· 00 ··• __ ·· __ •·••• __ ..··......... 51 10. New Tipulidae from the Society Islands, by Charles P. Alexander (1 figure) ...... 53 11. Demaptera and Orthoptera from the Society Islands, by Morgan Hebard (1 figure) __ 00 __ . __ __ . __ .. __ 57 12. Anthribidae from the Society Islands, by Karl Jordan (1 figure) 00 __ ..00........... 67 13. Dolichopodids from the Society Islands, by C. G. Lamb (1 plate) 71 14. Ceratopogonidae from the Society Islands, by J. w. S. Macfie (3 figures)............ 75 15. Staphylinidae from the Society Islands, by Malcolm Cameron (1 figure) 81 16. Some Tahitian Mycetophilidae and Chironomidae, by F. W. Edwards __ .. __ 85 17. Porcellio (Heminagara) Tahitiensis, new subgenus and species, and other Tahi- tian tesrrestrial Isopods, by Harold Gordon Jackson (1 figure) ....