Historical Range of Variation and State and Transition Modeling of Historic and Current Landscape Conditions for Potential Natural Vegetation Types of the Southwest
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Interior Arizona Chaparral
Rapid Assessment Reference Condition Model The Rapid Assessment is a component of the LANDFIRE project. Reference condition models for the Rapid Assessment were created through a series of expert workshops and a peer-review process in 2004 and 2005. For more information, please visit www.landfire.gov. Please direct questions to [email protected]. Potential Natural Vegetation Group (PNVG) R3CHAPsw Interior Arizona Chaparral General Information Contributors (additional contributors may be listed under "Model Evolution and Comments") Modelers Reviewers Tyson Swetnam [email protected] Linda Wadleigh [email protected] Reese Lolley [email protected] Vegetation Type General Model Sources Rapid AssessmentModel Zones Shrubland Literature California Pacific Northwest Local Data Great Basin South Central Dominant Species* Expert Estimate Great Lakes Southeast Northeast S. Appalachians QUTU LANDFIRE Mapping Zones CEGR Northern Plains Southwest 14 24 28 N-Cent.Rockies APPR 15 25 13 QUPU 23 27 Geographic Range Central and Northern Arizona, Central New Mexico. Some patches associated with Sky Islands of Southern Arizona and New Mexico. Also extends into the Mojave Desert and southern Great Basin. Biophysical Site Description Occurs across central Arizona (Mogollon Rim), and western New Mexico. It dominates along the mid- elevation transition from the Mojave, Sonoran, and Northern Chihuahuan deserts into mountains (1000- 2200 m). It occurs along foothills, mountain slopes and canyons in drier habitats below the encinal and Pinus Ponderosa woodlands. Stands are often associated with xeric coarse-textured substrates such as limestone, basalt or alluvium, especially in transition areas with more mesic woodlands (NatureServe 2004). Vegetation Description Vegetation is less dense than California chaparral, with aerial coverage of 35-80% ground surface in Arizona (Cable 1957, Carmichael et al. -

Adenostoma Sparsifolium Torr. (Rosaceae), Arctostaphylos Peninsularis Wells (Ericaceae), Artemisia Tridentata Nutt
66 JOURNAL OF THE LEPIDOPTERISTS' SOCIETY Ceanothus greggii A. Gray (Rhamnaceae), Adenostoma sparsifolium Torr. (Rosaceae), Arctostaphylos peninsularis Wells (Ericaceae), Artemisia tridentata Nutt. (Asteraceae), Quercus chrysolepis Liebm. and Q. dumosa Nutt. (Fagaceae), and Pinus jefferyi Grev. & BaH. (Pinaceae). On 27 and 29 October 1989 the unmated females were caged at a site in the vicinity of Mike's Sky Ranch in the Sierra San Pedro Martir, approximately 170 km south of the international border. Despite sunny weather and at a similar elevation and floral com munity, no males were attracted. Two males were deposited as voucher specimens in both of the following institutions: Universidad Autonoma de Baja California Norte, Ensenada, Mexico, and the Essig Mu seum of Entomology, University of California, Berkeley. Eleven specimens are in the private collection of John Noble, Anaheim Hills, California; the remaining 22 specimens are in the collection of the author. RALPH E. WELLS, 303-8 Hoffman Street, Jackson, California 95642. Received for publication 10 February 1990; revised and accepted 15 March 1991. Journal of the Lepidopterists' SOCiety 45(1), 1991, 66-67 POSITIVE RELATION BETWEEN BODY SIZE AND ALTITUDE OF CAPTURE SITE IN TORTRICID MOTHS (TORTRICIDAE) Additional key words: North America, biometrics, ecology. Earlier I reported a positive correlation between forewing length and altitude of capture site in the Nearctic tortricid Eucosma agricolana (Walsingham) (Miller, W. E. 1974, Ann. Entomol. Soc. Amer. 67:601-604). The all-male sample was transcontinental, with site altitudes ranging from near sea level on east and west coasts to more than 2700 m in the Rocky Mountains. Altitudes of capture came from labels of some specimens, and from topographic maps for others. -

Ceanothus Crassifolius Torrey NRCS CODE: Family: Rhamnaceae (CECR) Order: Rhamnales Subclass: Rosidae Class: Magnoliopsida
I. SPECIES Ceanothus crassifolius Torrey NRCS CODE: Family: Rhamnaceae (CECR) Order: Rhamnales Subclass: Rosidae Class: Magnoliopsida Lower right: Ripening fruits, two already dehisced. Lower center: Longitudinal channeling in stems of old specimen, typical of obligate seeding Ceanothus (>25 yr since last fire). Note dark hypanthium in center of white flowers. Photos by A. Montalvo. A. Subspecific taxa 1. C. crassifolius Torr. var. crassifolius 2. C. crassifolius Torr. var. planus Abrams (there is no NRCS code for this taxon) B. Synonyms 1. C. verrucosus Nuttal var. crassifolius K. Brandegee (Munz & Keck 1968; Burge et al. 2013) 2. C. crassifolius (in part, USDA PLANTS 2019) C. Common name 1. hoaryleaf ceanothus, sometimes called thickleaf ceanothus or thickleaf wild lilac (Painter 2016) 2. same as above; flat-leaf hoary ceanothus and flat-leaf snowball ceanothus are applied to other taxa (Painter 2016) D. Taxonomic relationships Ceanothus is a diverse genus with over 50 taxa that cluster in to two subgenera. C. crassifolius has long been recognized as part of the Cerastes group of Ceanothus based on morphology, life-history, and crossing studies (McMinn 1939a, Nobs 1963). In phylogenetic analyses based on RNA and chloroplast DNA, Hardig et al. (2000) found C. crassifolius clustered into the Cerastes group and in each analysis shared a clade with C. ophiochilus. In molecular and morphological analyses, Burge et al. (2011) also found C. crassifolius clustered into Cerastes. Cerastes included over 20 taxa and numerous subtaxa in both studies. Eight Cerastes taxa occur in southern California (see I. E. Related taxa in region). E. Related taxa in region In southern California, the related Cerastes taxa include: C. -

List of Approved Plants
APPENDIX "X" – PLANT LISTS Appendix "X" Contains Three (3) Plant Lists: X.1. List of Approved Indigenous Plants Allowed in any Landscape Zone. X.2. List of Approved Non-Indigenous Plants Allowed ONLY in the Private Zone or Semi-Private Zone. X.3. List of Prohibited Plants Prohibited for any location on a residential Lot. X.1. LIST OF APPROVED INDIGENOUS PLANTS. Approved Indigenous Plants may be used in any of the Landscape Zones on a residential lot. ONLY approved indigenous plants may be used in the Native Zone and the Revegetation Zone for those landscape areas located beyond the perimeter footprint of the home and site walls. The density, ratios, and mix of any added indigenous plant material should approximate those found in the general area of the native undisturbed desert. Refer to Section 8.4 and 8.5 of the Design Guidelines for an explanation and illustration of the Native Zone and the Revegetation Zone. For clarity, Approved Indigenous Plants are considered those plant species that are specifically indigenous and native to Desert Mountain. While there may be several other plants that are native to the upper Sonoran Desert, this list is specific to indigenous and native plants within Desert Mountain. X.1.1. Indigenous Trees: COMMON NAME BOTANICAL NAME Blue Palo Verde Parkinsonia florida Crucifixion Thorn Canotia holacantha Desert Hackberry Celtis pallida Desert Willow / Desert Catalpa Chilopsis linearis Foothills Palo Verde Parkinsonia microphylla Net Leaf Hackberry Celtis reticulata One-Seed Juniper Juniperus monosperma Velvet Mesquite / Native Mesquite Prosopis velutina (juliflora) X.1.2. Indigenous Shrubs: COMMON NAME BOTANICAL NAME Anderson Thornbush Lycium andersonii Barberry Berberis haematocarpa Bear Grass Nolina microcarpa Brittle Bush Encelia farinosa Page X - 1 Approved - February 24, 2020 Appendix X Landscape Guidelines Bursage + Ambrosia deltoidea + Canyon Ragweed Ambrosia ambrosioides Catclaw Acacia / Wait-a-Minute Bush Acacia greggii / Senegalia greggii Catclaw Mimosa Mimosa aculeaticarpa var. -

LANDFIRE Biophysical Setting Model Biophysical Setting 2611040 Mogollon Chaparral
LANDFIRE Biophysical Setting Model Biophysical Setting 2611040 Mogollon Chaparral This BPS is lumped with: This BPS is split into multiple models: General Information Contributors (also see the Comments field) Date 7/19/2005 Modeler 1 Matt Brooks [email protected] Reviewer Modeler 2 Reviewer Modeler 3 Reviewer Vegetation Type Dominant Species Map Zone Model Zone CAHO3 Upland Shrubland 26 Alaska Northern Plains ARPU5 California N-Cent.Rockies General Model Sources CEGR Great Basin Pacific Northwest Literature CEMO2 Great Lakes South Central Local Data QUTU2 Hawaii Southeast Expert Estimate PUST Northeast S. Appalachians QUPA4 Southwest Geographic Range This ecological system occurs across central AZ (Mogollon Rim), central to south central and western NM, southern UT, and eastern and southeastern NV (MZs 17, 13 and 14). It often dominates along the mid-elevation transition from the Mojave, Sonoran and northern Chihuahuan deserts. Biophysical Site Description This BpS is found in mountains from 1000-2200m. It occurs on foothills, mountain slopes and canyons in drier habitats below the encinal (southwestern oak woodlands) and Pinus ponderosa woodlands and above desert grasslands. Stands are often associated with more xeric and coarse-textured substrates such as limestone, basalt or alluvium, especially in transition areas with more mesic woodlands. Typical of xeric montane moderate to steep slopes. The soils are generally shallow and derived from granite, basalt or other igneous volcanic rocks. These soils occur in mesic (moderately warm) -

Reference Plant List
APPENDIX J NATIVE & INVASIVE PLANT LIST The following tables capture the referenced plants, native and invasive species, found throughout this document. The Wildlife Action Plan Team elected to only use common names for plants to improve the readability, particular for the general reader. However, common names can create confusion for a variety of reasons. Common names can change from region-to-region; one common name can refer to more than one species; and common names have a way of changing over time. For example, there are two widespread species of greasewood in Nevada, and numerous species of sagebrush. In everyday conversation generic common names usually work well. But if you are considering management activities, landscape restoration or the habitat needs of a particular wildlife species, the need to differentiate between plant species and even subspecies suddenly takes on critical importance. This appendix provides the reader with a cross reference between the common plant names used in this document’s text, and the scientific names that link common names to the precise species to which writers referenced. With regards to invasive plants, all species listed under the Nevada Revised Statute 555 (NRS 555) as a “Noxious Weed” will be notated, within the larger table, as such. A noxious weed is a plant that has been designated by the state as a “species of plant which is, or is likely to be, detrimental or destructive and difficult to control or eradicate” (NRS 555.05). To assist the reader, we also included a separate table detailing the noxious weeds, category level (A, B, or C), and the typical habitats that these species invade. -

Transactions
DESERT BIGHORN COUNCIL TRANSACTIONS VOLUME 45 Desert Bighorn Council 2001 Transactions A Compilation of Papers Presented at the 45thAnnual Meeting Hermosillo, Sonora, Mexico April 18-21,2001 Jonathan D. Hanna, Editor Arizona Game and Fish Department 7200 E. University, Mesa, AZ 85207 Reviewers for the 200 1 Desert Bighorn Council Transactions: Francisco Abarca, Clay Brewer, Bob Henry, Dana McGehee, Ted McKinney, Eric Rominger Illustrations by Pat Hansen Carlos Castdo and Jim debs Secretary Darren Divine Treasurer Charles Douglas William Brighanl (Chair), James DeForge, Amy Fisher, Mark Jorgensen, Raymond Lee, and John W ehausen Richard Weaver Mara W eisenberger Gary Montoya Trans actions : Raymond Lee Ross Haley Warren Kelly Doris Weaver William Brigham Special thanks to the Foundation for North herican Wild Sheep for their generous support of the Desert Bighorn Cow~cilAnnual Meeting Brian F. Wakeling ... ............................................................................ .2 Brian F. Wakeling ...........................................................................................................................4 Eric M. Rorninger and Elise Goldstein ....................................................................................... .6 ESTATUS DEL GO CHMAIRlEaON EN NEW EXIC8 EN EL 2008. Eric M. Romhger y Elise Goldstein ............................................................................................ -13 $ OF DESERT BPGXOW SHEEP PlFJ TEXAS - 2008. Michael T. Bitt hael D. Hobson ................................................. -
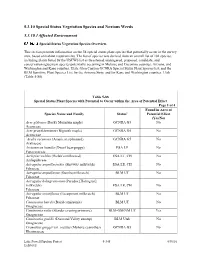
Special Status Vegetation Species and Noxious Weeds
5.3.10 Special Status Vegetation Species and Noxious Weeds 5.3.10.1 Affected Environment Special Status Vegetation Species Overview. This section presents information on the 58 special status plant species that potentially occur in the survey area, based on habitat requirements. The list of species was derived from an overall list of 101 species, including plants listed by the USFWS list as threatened, endangered, proposed, candidate, and conservation agreement species potentially occurring in Mohave and Coconino counties, Arizona, and Washington and Kane counties, Utah; Glen Canyon GCNRA Special Status Plant Species list; and the BLM Sensitive Plant Species List for the Arizona Strip, and for Kane and Washington counties, Utah (Table 5-88). Table 5-88 Special Status Plant Species with Potential to Occur within the Area of Potential Effect Page 1 of 4 Found in Area of Species Name and Family Status1 Potential Effect (Yes/No) Acer glabrum (Rocky Mountain maple) GCNRA G5 No Aceraceae Acer grandidentatum (Bigtooth maple) GCNRA G5 No Aceraceae `Aralia racemosa (American spikenard) GCNRA G5 No Araliaceae Arctomecon humilis (Dwarf bear-poppy) ESA LE No Papaveraceae Asclepias welshii (Welsh’s milkweed) ESA LT, CH No Asclepidaceae Astragalus ampullarioides (Shivwits milkvetch) ESA LE, CH No Fabaceae Astragalus ampullarius (Gumbo milkvetch) BLM UT No Fabaceae Astragalus holmgreniorum (Paradox [Holmgren] milkvetch) ESA LE, CH No Fabaceae Astragalus striatiflorus (Escarpment milkvetch) BLM UT No Fabaceae Camissonia bairdii (Baird camissonia) BLM -

Inventory of Sensitive Species and Ecosystems in Utah, Endemic And
,19(1725<2)6(16,7,9(63(&,(6$1'(&26<67(06 ,187$+ (1'(0,&$1'5$5(3/$1762)87$+ $129(59,(:2)7+(,5',675,%87,21$1'67$786 &HQWUDO8WDK3URMHFW&RPSOHWLRQ$FW 7LWOH,,,6HFWLRQ E 8WDK5HFODPDWLRQ0LWLJDWLRQDQG&RQVHUYDWLRQ&RPPLVVLRQ 0LWLJDWLRQDQG&RQVHUYDWLRQ3ODQ 0D\ &KDSWHU3DJH &RRSHUDWLYH$JUHHPHQW1R8& 6HFWLRQ9$D 3UHSDUHGIRU 87$+5(&/$0$7,210,7,*$7,21$1'&216(59$7,21&200,66,21 DQGWKH 86'(3$570(172)7+(,17(5,25 3UHSDUHGE\ 87$+',9,6,212):,/'/,)(5(6285&(6 -81( 6#$.'1(%106'065 2CIG #%-019.'&)/'065 XKK +0641&7%6+10 9*;&1'576#**#8'51/#0;4#4'2.#065! 4CTKV[D['EQTGIKQP 4CTKV[D[5QKN6[RG 4CTKV[D[*CDKVCV6[RG 'PFGOKEUD[.KHG(QTO 'PFGOKEUD[#IGCPF1TKIKP *+5614;1(4#4'2.#06+08'0614;+076#* 75(KUJCPF9KNFNKHG5GTXKEG 1VJGT(GFGTCN#IGPEKGU 7VCJ0CVKXG2NCPV5QEKGV[ 7VCJ0CVWTCN*GTKVCIG2TQITCO *196175'6*+54'2146 $CUKUHQT+PENWUKQP 0QVGUQP2NCPV0QOGPENCVWTG 5VCVWU%CVGIQTKGUHQT+PENWFGF2NCPVU )GQITCRJKE&KUVTKDWVKQP 2NCPVUD[(COKN[ #TGCUHQT#FFKVKQPCN4GUGCTEJ '0&'/+%#0&4#4'2.#0651(76#* *KUVQTKECN 4CTG 9CVEJ 2GTKRJGTCN +PHTGSWGPV 6CZQPQOKE2TQDNGOU #FFKVKQPCN&CVC0GGFGF KKK .+6'4#674'%+6'& $SSHQGL[$ 3ODQWVZLWK)HGHUDO$JHQF\6WDWXV $SSHQGL[% 3ODQWVE\&RXQW\ $SSHQGL[& 3ODQWVE\)DPLO\ +0&': KX .+561(6#$.'5 2CIG 6CDNG *KUVQT[QH7VCJRNCPVVCZCNKUVGFQTTGXKGYGFCUECPFKFCVGU HQTRQUUKDNGGPFCPIGTGFQTVJTGCVGPGFNKUVKPIWPFGTVJG HGFGTCN'PFCPIGTGF5RGEKGU#EV 6CDNG 0WOGTKECNCPCN[UKUQHRNCPVVCZCD[UVCVWUECVGIQT[ .+561((+)74'5 (KIWTG 'EQTGIKQPUQHVJGYGUVGTP7PKVGF5VCVGU X XK $&.12:/('*0(176 7KLVUHYLHZRIHQGHPLFDQGUDUHSODQWVSHFLHVLVDFRPSRQHQWRIDODUJHULQWHUDJHQF\HIIRUWWR FRPSOHWHDQLQYHQWRU\RIVHQVLWLYHVSHFLHVDQGHFRV\VWHPVLQ8WDK7KH8WDK'LYLVLRQRI:LOGOLIH -

Plant Palette for the Town of Camp Verde, Arizona a Tree City USA Since 2014
Suggested Plant Palette For the Town of Camp Verde, Arizona A Tree City USA Since 2014 Second Edition 2020 Compiled by The Camp Verde Tree Advisory Committee Plant Palette for the Town of Camp Verde Second Edition- 2020 Plant Palette for the Town of Camp Verde Second Edition- 2020 Table of Contents INTRODUCTION THE RIGHT PLANT IN THE RIGHT PLACE FLOWERS 2 ANGELITA DAISY (TETRANEURIS ACAULIS) 3 BABY'S BREATH (GYSOPHILA PANICULATA) 4 CHOCOLATE FLOWER (BERLANDERIA LYRATA ) 5 DESERT GLOBE MALLOW (SPHAERALCEA AMBIGUA) 6 DESERT MARIGOLD (BAILEYA MULTIRADIATA) 7 FIREWHEEL (GAILLARDIA PULCHELLA) 8 GOLDEN COLUMBINE (AQUILEGIA CHRYSANTHA) 9 GOODDING VERBENA (GLANDULARIA GOODINGII) 10 MEXICAN EVENING PRIMROSE (OENOTHER SPECIOSA) 11 FIRECRACKER PENSTEMON (PENSTEMON EATONII) 12 PALMER’S PENSTEMON (PENSTEMON PALMERI) 12 PARRY’S PENSTEMON (PENSTEMON PARRYI) 13 PURPLE CONE FLOWER (ECHINACEA PURPUREA) 14 RED HOT POKER (KNOPHIA SSP.) 15 ROSEMARY (ROSMARINUS OFFICINALIS 'PROSTRATUS') 16 SANDPAPER VERBENA (VERBENA RIGIDA) 17 SHOWY MILKWEED (ASCLEPIAS SPECIOSA) 18 GRASSES 19 BEARGRASS (NOLINA MICROCARPA) 20 BLUE GRAMA GRASS (BOUTELOUA GRACILIS) 21 BUFFALOGRASS (BOUTELOUA DACTYLOIDES) 22 DEER GRASS (MULENBERGIA RIGENS) 23 LITTLE BLUESTEM (SCHIZACHYRIUM SCOPARIUM) 24 PINK MUHLYGRASS (MULENBERGIA CAPILLARIS) 25 PURPLE THREE AWN (ARISTIDA PURPUREA) 26 SHRUBS 27 AUTUMN SAGE (SALVIA GREGGII) 28 BUSH DALEA/INDIGO BUSH (DALEA PULCHRA) 29 CHIHUAHUAN SAGE (LEUCOPHYLLUM LAEVIGATUM) 30 DESERT CEANOTHUS (CEANOTHUS GREGGII A. GRAY) 31 DESERT HONEYSUCKLE (ANISACANTHUS THURBERI) -

Vegetation Alliances of the San Dieguito River Park Region, San Diego County, California
Vegetation alliances of the San Dieguito River Park region, San Diego County, California By Julie Evens and Sau San California Native Plant Society 2707 K Street, Suite 1 Sacramento CA, 95816 In cooperation with the California Natural Heritage Program of the California Department of Fish and Game And San Diego Chapter of the California Native Plant Society Final Report August 2005 TABLE OF CONTENTS Introduction...................................................................................................................................... 1 Methods ........................................................................................................................................... 2 Study area ................................................................................................................................... 2 Existing Literature Review........................................................................................................... 2 Sampling ..................................................................................................................................... 2 Figure 1. Study area including the San Dieguito River Park boundary within the ecological subsections color map and within the County inset map............................................................ 3 Figure 2. Locations of the field surveys....................................................................................... 5 Cluster analyses for vegetation classification ............................................................................ -

Moth Larval Host Plants — Desert Survivors (Dec. 2017) Scientific Name Common Name Caterpillars That Use These Species Abutilon Spp
Moth Larval Host Plants — Desert Survivors (Dec. 2017) Scientific Name Common Name Caterpillars that Use These Species Abutilon spp. Desert mallow Bird dropping moth, Owlet moth, Crambid seed moth Acaciella hirta (Acacia Fernball acacia Silk moth, Mesquite stinger flannel angustissima) moth Agastache wrightii Wright’s horsemint Crambid moth Agave spp. Century plant, agave Yucca moth, Tortrix moth Aloysia wrightii Wright's beebrush Rustic sphinx moth Ambrosia spp. Ragweed Bird dropping moth, Geometrid moth Amorpha spp. False indigo bush Tortrix leaf roller moth, Pyralid moth Artemisia ludoviciana subsp. Western mugwort, white Flower moth, Tortrix moth redolens sagebrush Asclepias spp. Milkweed Milkweed tussock tiger moth Atriplex spp. Saltbush Geometrid moth Baccharis brachyphylla Shortleaf baccharis Owlet moth Baccharis salicifolia Seep willow Tiger moth Baccharis sarathroides Desert broom Flower moth, Owlet moth, Geometrid moth, Desert Broom gall moth Bahia absinthifolia Desert bahia Owlet moth Bebbia juncea var. aspera Sweet bush Tiger moth, Tortrix moth Bidens aurea Arizona beggarticks Sunflower moth, Dart moth Bothriochloa barbinodis Cane beardgrass Owlet moth Bouteloua curtipendula Sideoats grama Hualapai buckmoth, Viened Ctenucha moth Bouvardia ternifolia Smooth bouvardia Falco sphinx moth Brickellia californica California brickelbush Flower moth, Geometrid moth, Northern giant tiger moth Buddleja sessiliflora Rio Grande butterfly bush Hooded owlet moth Calliandra eriophylla Native fairyduster (2) Melipotis moths Ceanothus greggii Desert deerbrush Cecrops eyed silkmoth, Prominent moth, (4) geometrid moths, Owlet moth Celtis pallida Desert hackberry Small prominent moth, Randa’s eyed silk moth Celtis reticulata Canyon hackberry Io Silkmoth, Small prominent moth, Cleft headed looper moth !1 Scientific Name Common Name Caterpillars that Use These Species Calylophus hartwegii Sundrops White lined sphinx moth, Flower moth Cercocarpus breviflorus Hairy mountain mahogany Cecrops eyed silkmoth, Owlet moth, (4) Geometrid moths, Sulphur moth Chilopsis linearis subsp.