Shrublands in California: Literature Review and Research Needed for Management
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Palm Haven Garden Plant List - 2010
PALM HAVEN GARDEN PLANT LIST - 2010 BOTANICAL NAME COMMON NAME Achillea millefolium 'Paprika' Paprika yarrow Arctostaphylos glauca big berry manzanita Arctostaphylos pajaroensis Pajaro manzanita Arctostaphylos uva-ursi bearberry Arctostaphylos uva-ursi 'San Bruno Mountain' San Bruno Mountain bearberry Aristolochia californica California pipevine Artemisia pycnocephala coastal sagewort Baccharis pilularis 'Twin Peaks' Twin Peaks dwarf coyote brush Berberis aquifolium 'Golden Abundance' Golden Abundance Oregon-grape Carex spissa San Diego sedge Ceanothus 'Joyce Coulter' Joyce Coulte wild lilac Ceanothus arboreus feltleaf ceanothus Ceanothus gloriosus var exaltatus 'Emily Brown' Emily Brown glory brush Dendromecon harfordii Channel Island tree poppy Epilobium canum California fuchsia Erigeron glaucus seaside daisy Eriogonum fasciculatum California buckwheat Eriogonum latifolium coast buckwheat Eriogonum umbellatum sulfur flower Eriophyllum confertiflorum golden yarrow Heteromeles arbutifolia toyon Heterotheca sessilifolora false goldenaster Iris douglasiana Douglas iris Leymus condensatus 'Canyon Prince' Canyon Prince giant wild rye Mimulus aurantiacus sticky monkeyflower Monardella villosa coyote mint Mulhenbergia rigens deer grass Penstemon heterophyllus foothill penstemon Physocarpus capitatus ninebark Rhamnus californica 'Ed Holm' Ed Holm coffeeberry Rhamnus californica 'Mound San Bruno' Mound San Bruno coffeeberry Salvia clevelandii Cleveland sage Salvia mellifera 'Shirley's Creeper' Shirley's Creepe sage Salvia sonomensis 'Bee's Bliss' Bee's Bliss sage Satureja douglasii yerba buena Trichostema lanatum wolly blue curls Typha angustifolia narrow-leaved cattail Verbena lilacina lilac verbena Vitis californica 'Roger's Red' Roger's Red California wild grape. -

April 26, 2019
April 26, 2019 Theodore Payne Foundation’s Wild Flower Hotline is made possible by donations, memberships, and the generous support of S&S Seeds. Now is the time to really get out and hike the trails searching for late bloomers. It’s always good to call or check the location’s website if you can, and adjust your expectations accordingly before heading out. Please enjoy your outing, and please use your best flower viewing etiquette. Along Salt Creek near the southern entrance to Sequoia National Park, the wildflowers are abundant and showy. Masses of spring flowering common madia (Madia elegans) are covering sunny slopes and bird’s-eye gilia (Gilia tricolor) is abundant on flatlands. Good crops of owl’s clover (Castilleja sp.) are common in scattered colonies and along shadier trails, woodland star flower (Lithophragma sp.), Munz’s iris (Iris munzii), and the elegant naked broomrape (Orobanche uniflora) are blooming. There is an abundance of Chinese houses (Collinsia heterophylla) and foothill sunburst (Pseudobahia heermanii). This is a banner year for the local geophytes. Mountain pretty face (Tritelia ixiodes ssp. anilina) and Ithuriel’s spear (Triteliea laxa) are abundant. With the warming temperatures farewell to spring (Clarkia cylindrical subsp. clavicarpa) is starting to show up with their lovely bright purple pink floral display and is particularly noticeable along highway 198. Naked broom rape (Orobanche uniflora), foothill sunburst (Pseudobahia heermanii). Photos by Michael Wall © Theodore Payne Foundation for Wild Flowers & Native Plants, Inc. No reproduction of any kind without written permission. The trails in Pinnacles National Park have their own personality reflecting the unusual blooms found along them. -

The Vegetation of Robinson Crusoe Island (Isla Masatierra), Juan
The Vegetation ofRobinson Crusoe Island (Isla Masatierra), Juan Fernandez Archipelago, Chile1 Josef Greimler,2,3 Patricio Lopez 5., 4 Tod F. Stuessy, 2and Thomas Dirnbiick5 Abstract: Robinson Crusoe Island of the Juan Fernandez Archipelago, as is the case with many oceanic islands, has experienced strong human disturbances through exploitation ofresources and introduction of alien biota. To understand these impacts and for purposes of diversity and resource management, an accu rate assessment of the composition and structure of plant communities was made. We analyzed the vegetation with 106 releves (vegetation records) and subsequent Twinspan ordination and produced a detailed colored map at 1: 30,000. The resultant map units are (1) endemic upper montane forest, (2) endemic lower montane forest, (3) Ugni molinae shrubland, (4) Rubus ulmifolius Aristotelia chilensis shrubland, (5) fern assemblages, (6) Libertia chilensis assem blage, (7) Acaena argentea assemblage, (8) native grassland, (9) weed assemblages, (10) tall ruderals, and (11) cultivated Eucalyptus, Cupressus, and Pinus. Mosaic patterns consisting of several communities are recognized as mixed units: (12) combined upper and lower montane endemic forest with aliens, (13) scattered native vegetation among rocks at higher elevations, (14) scattered grassland and weeds among rocks at lower elevations, and (15) grassland with Acaena argentea. Two categories are included that are not vegetation units: (16) rocks and eroded areas, and (17) settlement and airfield. Endemic forests at lower elevations and in drier zones of the island are under strong pressure from three woody species, Aristotelia chilensis, Rubus ulmifolius, and Ugni molinae. The latter invades native forests by ascending dry slopes and ridges. -
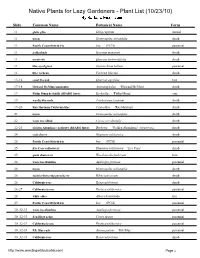
Native Plants for Lazy Gardeners - Plant List (10/23/10)
Native Plants for Lazy Gardeners - Plant List (10/23/10) Slide Common Name Botanical Name Form 11 globe gilia Gilia capitata annual 11 toyon Heteromeles arbutifolia shrub 11 Pacific Coast Hybrid iris Iris (PCH) perennial 11 goldenbush Isocoma menziesii shrub 11 scrub oak Quercus berberidifolia shrub 11 blue-eyed grass Sisyrinchium bellum perennial 11 lilac verbena Verbena lilacina shrub 13-16 coast live oak Quercus agrifolia tree 17-18 Howard McMinn man anita Arctostaphylos 'Howard McMinn' shrub 19 Philip Mun keckiella (RSABG Intro) Keckiella 'Philip Munz' ine 19 woolly bluecurls Trichostema lanatum shrub 19-20 Ray Hartman California lilac Ceanothus 'Ray Hartman' shrub 21 toyon Heteromeles arbutifolia shrub 22 western redbud Cercis occidentalis shrub 22-23 Golden Abundance barberry (RSABG Intro) Berberis 'Golden Abundance' (MAHONIA) shrub 2, coffeeberry Rhamnus californica shrub 25 Pacific Coast Hybrid iris Iris (PCH) perennial 25 Eve Case coffeeberry Rhamnus californica '. e Case' shrub 25 giant chain fern Woodwardia fimbriata fern 26 western columbine Aquilegia formosa perennial 26 toyon Heteromeles arbutifolia shrub 26 fuchsia-flowering gooseberry Ribes speciosum shrub 26 California rose Rosa californica shrub 26-27 California fescue Festuca californica perennial 28 white alder Alnus rhombifolia tree 29 Pacific Coast Hybrid iris Iris (PCH) perennial 30 032-33 western columbine Aquilegia formosa perennial 30 032-33 San Diego sedge Carex spissa perennial 30 032-33 California fescue Festuca californica perennial 30 032-33 Elk Blue rush Juncus patens '.l1 2lue' perennial 30 032-33 California rose Rosa californica shrub http://www weedingwildsuburbia com/ Page 1 30 032-3, toyon Heteromeles arbutifolia shrub 30 032-3, fuchsia-flowering gooseberry Ribes speciosum shrub 30 032-3, Claremont pink-flowering currant (RSA Intro) Ribes sanguineum ar. -

Interior Arizona Chaparral
Rapid Assessment Reference Condition Model The Rapid Assessment is a component of the LANDFIRE project. Reference condition models for the Rapid Assessment were created through a series of expert workshops and a peer-review process in 2004 and 2005. For more information, please visit www.landfire.gov. Please direct questions to [email protected]. Potential Natural Vegetation Group (PNVG) R3CHAPsw Interior Arizona Chaparral General Information Contributors (additional contributors may be listed under "Model Evolution and Comments") Modelers Reviewers Tyson Swetnam [email protected] Linda Wadleigh [email protected] Reese Lolley [email protected] Vegetation Type General Model Sources Rapid AssessmentModel Zones Shrubland Literature California Pacific Northwest Local Data Great Basin South Central Dominant Species* Expert Estimate Great Lakes Southeast Northeast S. Appalachians QUTU LANDFIRE Mapping Zones CEGR Northern Plains Southwest 14 24 28 N-Cent.Rockies APPR 15 25 13 QUPU 23 27 Geographic Range Central and Northern Arizona, Central New Mexico. Some patches associated with Sky Islands of Southern Arizona and New Mexico. Also extends into the Mojave Desert and southern Great Basin. Biophysical Site Description Occurs across central Arizona (Mogollon Rim), and western New Mexico. It dominates along the mid- elevation transition from the Mojave, Sonoran, and Northern Chihuahuan deserts into mountains (1000- 2200 m). It occurs along foothills, mountain slopes and canyons in drier habitats below the encinal and Pinus Ponderosa woodlands. Stands are often associated with xeric coarse-textured substrates such as limestone, basalt or alluvium, especially in transition areas with more mesic woodlands (NatureServe 2004). Vegetation Description Vegetation is less dense than California chaparral, with aerial coverage of 35-80% ground surface in Arizona (Cable 1957, Carmichael et al. -

Flora of the Mediterranean Basin in the Chilean Espinales: Evidence of Colonisation
PASTOS 2012. ISSN: 0210-1270 PASTOS, 42 (2), 137 - 160 137 FLORA OF THE MEDITERRANEAN BASIN IN THE CHILEAN ESPINALES: EVIDENCE OF COLONISATION I. MARTÍN-FORÉS1, M. A. CASADO1*, I. CASTRO2, C. OVALLE3, A. DEL POZO4, B. ACOSTA-GALLO1, L. SÁNCHEZ-JARDÓN1 AND J. M. DE MIGUEL1 1Departamento de Ecología. Facultad de Biología. Universidad Complutense de Madrid. Madrid (España). 2Departamento de Ecología. Facultad de Ciencias. Universidad Autónoma de Madrid. Madrid (España). 3Instituto de Investigaciones Agropecuarias INIA-La Cruz. La Cruz (Chile). 4Facultad de Ciencias Agrarias. Universidad de Talca. Talca (Chile). *Author for correspondence: M.A. Casado ([email protected]). SUMMARY In Chile’s Mediterranean region, over 18% of plant species are alien. This is particularly noteworthy in some agrosilvopastoral systems such as the espinales, which are functionally very similar to the Spanish dehesas and are of great ecological and socioeconomic interest. In the present paper we analyse Chile’s non-native flora, considering three scales of analysis: national, regional (the central region, presenting a Mediterranean climate) and at community level (the espinales within the central region). We compare this flora with that recorded in areas of the Iberian Peninsula with similar lithological and geomorphological characteristics, and land use. We discuss possible mechanisms that might have been operating in the floristic colonisation from the Mediterranean Basin to Chile’s Mediterranean region. Key words: Alien plants, biogeography, Chile, life cycle, Spain. INTRODUCTION Historically, the transit of goods, domestic animals and people has favoured the flow of wild organisms around the planet (Lodge et al., 2006), a fact that is often associated with the introduction of cultural systems, which have contributed to generating new environmental and socioeconomic scenarios (Le Houérou, 1981; Hobbs, 1998; Grenon and Batisse, 1989). -

Piedra Blanca Trail Middle Sespe Creek/Pine Mountain Ridge, Ventura County, California by David L
Vascular Plants of the Piedra Blanca Trail Middle Sespe Creek/Pine Mountain Ridge, Ventura County, California By David L. Magney Botanical Name Common Name Habit Family Acer macrophyllum Bigleaf Maple T Sapindaceae Acmispon ? Lotus AH Fabaceae Acmispon glaber var. glaber Deerweed S Fabaceae Acmispon strigosus var. strigosus Strigose Lotus AH Fabaceae Acourtia microcephala Sacapellote PH Asteraceae Adenostoma fasciculatum Chamise S Rosaceae Agoseris ? Mountain Dandelion PH Asteraceae Alnus rhombifolia White Alder T Betulaceae Amorpha californica False Indigo S Fabaceae Antirrhinum multiflorum Sticky Snapdragon S Veronicaceae Aquilegia formosa Columbine PH Ranunculaceae Arctostaphylos glauca Bigberry Manzanita S Ericaceae Artemisia douglasiana Mugwort S Asteraceae Artemisia tridentata ssp. tridentata Great Basin Sagebrush S Asteraceae Asclepias eriocarpa Woolly Milkweed AH Apocynaceae Astragalus ? Milkvetch AH Fabaceae Avena barbata* Slender Wild Oat AG Poaceae Baccharis salicifolia Mulefat S Asteraceae Boechera arcuata Few-flowered Rock Cress PH Brassicaceae Brickellia californica California Brickellbush S Asteraceae Bromus ? Brome PG Poaceae Bromus madritensis ssp. rubens* Red Brome AG Poaceae Bromus tectorum var. tectorum* Downy Brome AG Poaceae Calocedrus decurrens Incense-cedar T Cupressaceae Calyptridium monandrum Common Calyptridium AH Montiaceae Calystegia malacophylla ssp. cf pedicellata Sierra Morning-glory PH Convolvulaceae Camissonia boothii ssp. decorticans Shreading Evening Primrose AH Onagraceae Camissonia campestris ssp. campestris? Mojave Sun-cup AH Onagraceae Camissoniopsis micrantha Tiny Primrose AH Onagraceae Camissoniopsis pallida ssp. pallida Pale Primrose AH Onagraceae Carex ? Sedge PG Cyperaceae Carex senta Rough Sedge PG Cyperaceae Castilleja ? Indian Paintbrush PH Orobanchaceae Castilleja affinis ssp. affinis Lay-and-Collie's Indian Paintbrush PH Orobanchaceae Castilleja foliolosa Woolly Indian Paintbrush PH Orobanchaceae Castilleja subinclusa ssp. subinclusa Long-leaved Indian Paintbrush PH Orobanchaceae Caulanthus coulteri var. -

DISCUSSION of the FLORA of GUADALUPE ISLAND Dr. Reid Moran1
DISCUSSION OF THE FLORA OF GUADALUPE ISLAND Dr. Reid Moran1: Guadalupe Island lies about 250 miles south- southwest of San Diego, California, and about 160 miles off the peninsula of Baja California, Mexico. Volcanic in origin and sep arated from the peninsula by depths of about 12,000 feet, it is clearly an oceanic island. Among the vascular plants recorded from Guadalupe Island and its islets, apparently 164 species are native. Goats, introduced more than a century ago, have eliminated some species and re duced others nearly to extinction. Outer Islet, a goatless refugium two miles south of the main island, has a native florula of 36 species. Nine of these (including Euphorbia misera and Lavatera occidentalis) are very scarce on the main island, largely confined to cliffs inaccessible to goats; another one (Coreopsis gigantea) has not been collected there since 1875; and five others (includ ing Lavatera lindsayi, Dudleya guadalupensis, and Rhus integri¬ folia) have never been recorded from there. Although it is not known that these five ever did occur on the main island, presum ably they did but were exterminated by the goats. These five, comprising 14 per cent of the native florula of Outer Islet, give the only suggestion we have as to how many species must have been eliminated from the main island by the goats before they could be found by botanists. Also reported from Guadalupe Island are 42 species that prob ably are not native. Several of these, each found only once, ap parently have not persisted; but, with the severe reduction of many native plants by the goats, other introduced plants have become abundant. -

California Indian Garden – Plant List and Plant Uses
California Indian Garden Plant List Spring 2018 Common name Scientific name Indian Uses Bladderpod Isomeris arborea Seeds and flowers eaten Black sage Salvia mellifera Seeds ground into a meal for baking; tea made from leaves and stem Blue elderberry Sambucus nigra Berries used as food and sauce, plant also used for medicine, dyes for basketry, arrow shafts, flute, whistles, clapper sticks, and folk medicine Bush monkey flower Mimulus aurantiacus Young stems and leaves eaten as greens; used to treat burns, wounds, colds, cough, flu, stomach disorders and heart ailments CA buckwheat Eriogonum fasciculatum Leaf tea used for headache and stomach pain, root tea for colds and laryngitis; root poultice applied to wounds Chamise Adenostoma fasciculatum Infusion of bark and leaves used to cure syphilis; oils used to treat skin infections; scale insect on plant used as a binding agent; branches used to make arrow shafts and points Coast cholla Cylindropuntia prolifera Flowers and fruits for food Coast live oak Quercus agrifolia Acorns used as an important food staple Coast prickly pear Opuntia littoralis Fruit used for food, syrup, juice, candy and gum; young, green nopales (stems) also eaten; used to treat wounds, rheumatism, mumps, and reduce swelling; spines for needles and juice for dye Coast sunflower Encelia californica No known uses Coastal sagebrush Artemisia californica Leaves used for a variety of medicinal treatments: toothaches, wounds, asthma, colds, coughs, rheumatism, menstrual problems, to ease childbirth, menopausal symptoms, -

Contra Costa County, California
APPENDIX G BIOLOGICAL RESOURCES ASSESSMENT AND ARBORIST REPORTS Biological Resources Assessment for the Sufi Church Project Contra Costa County, California Prepared for: Meher Schools G-1 Prepared for: Meher Schools 999 Leland Drive Lafayette, CA 94549 925-938-9958 Prepared by: EDAW 2099 Mt. Diablo Blvd., Suite 204 Walnut Creek, CA 94596 (925) 279-0580 June 18, 2008 BIOLOGICAL RESOURCES ASSESSMENT FOR THE PROPOSED SUFI CHURCH PROJECT, CONTRA COSTA COUNTY, CALIFORNIA G-2 The information provided in this document is intended solely for the use and benefit of Meher Schools. No other person or entity shall be entitled to rely on the services, opinions, recommendations, plans or specifications provided herein, without the express written consent of EDAW, 2099 Mt. Diablo Blvd., Suite 204, Walnut Creek, CA 94596. G-3 TABLE OF CONTENTS SUMMARY.............................................................................................................................. i 1.0 INTRODUCTION AND METHODS .............................................................................1 2.0 EXISTING CONDITIONS.............................................................................................5 2.1 SETTING......................................................................................................................5 2.2 PLANT COMMUNITIES AND WILDLIFE HABITATS........................................................5 3.0 SPECIAL-STATUS BIOLOGICAL RESOURCES.......................................................7 3.1 SPECIAL-STATUS PLANTS ...........................................................................................7 -

A Self--Guided Tour
SONOMA STATE UNIVERSITY A SELF--GUIDED TOUR School of Social Sciences SONOMA- Department of Environmental Studies and Planning STATE UNIVERSITY . A self-guided tour Written by: Kenneth M. Stocking Professor ·Emeritus, Environmental Studies and Planning Robert J. Sherman, Professor of Biology Karen Tillinghast, Lead Gardener, Landscape Services 1st Revision, 1997, by Brian King and Karen Tillinghast 2nd Revision, 2006, by Katherine Musick and Karen Tillinghast ACKNOWLEDGEMENTS TABLE OF CONTENTS Our appreciation and thanks to the students, staff, and faculty from Environmental Studies and Planning and Biology who have contributed in many ways toward the development of the garden. Introduction ....................................................... iv Concept of the Garden . ...................... iv Special thanks to the following: Layout of the Garden. v John Bond, Director of Plant Operations, Retired A. Oak Woodland. 1 William Mabry, Director of Plant Operations, Retired B. Yellow Pine Forest ............. ................................. 4 George Smith, Superintendent of Grounds, Retired C. Douglas-Fir Forest .................... ...... ..................... 7 Sam Youney, Superintendent of Grounds D. Mixed Evergreen. 8 E. Grassland ...................................................... 10 Organizations that have contributed time, effort, and plant specimens include: F. Chaparral . 12 California Flora Nursery G. Redwood Forest ................................................ 15 California Native Plant Society, Milo Baker Chapter H. -

Vascular Plants Ventana Double Cone Trail
CALIFORNIA NATIVE PLANT SOCIETY – VASCULAR PLANTS VENTANA DOUBLE CONE TRAIL Acer macrophyllum - big-leaved maple Epilobium minutum - minute willow-herb Acmispon argophyllus - silver-leaved lotus Ericameria nauseosus var. speciosa - common rabbit-brush Acmispon glaber - deerweed Erigeron petrophilus - rock daisy Acmispon grandiflorus - large-flowered lotus Eriodictyon californicum - yerba santa Acmispon parviflorus - small-flowered lotus Eriogonum fasciculatum var. foliolosum - California buckwheat Adenostoma fasciculatum - chamise Eriogonum nudum var. pubiflorum - naked eriogonum/tibinagua Agoseris grandiflora - large-flowered agoseris Eriogonum saxatile - rock buckwheat Allophyllum gilioides - straggling gilia Eriophyllum confertiflorum - golden yarrow Antirrhinum multiflorum - sticky snapdragon Festuca microstachys - Nuttall's fescue Arbutus menziesii - madrone Festuca myuros - rattail fescue Arceuthobium campylopodum - western dwarf mistletoe Frangula californica - California coffeeberry Arctostaphylos glandulosa - Eastwood's manzanita Galium angustifolium - narrow-leaved bedstraw Boechera breweri - Brewer's rock cress Galium aparine - goose-grass Bromus carinatus var. carinatus - California brome Galium californicum ssp. flaccidum - California bedstraw Bromus diandrus - ripgut grass Galium californicum ssp. luciense - Lucia bedstraw Bromus grandis - tall brome Galium clementis - Santa Lucia bedstraw Bromus laevipes - woodland brome Galium porrigens - climbing bedstraw Bromus madritensis ssp. rubens - red brome Garrya flavescens - ashy