Research Article Morphological and Molecular Observation to Confirm
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Interaction of Drought and the Outbreak of Phoracantha
The interaction of drought and the outbreak of Phoracantha semipunctata (Coleoptera: Cerambycidae) on tree collapse in the Northern Jarrah (Eucalyptus marginata) forest. by Stephen Seaton (BSc Environmental Science) This thesis is presented in partial fulfilment of the requirements for the degree of Bachelor of Science (Honours) School of Biological Sciences and Biotechnology, Murdoch University, Perth, Western Australia November 2012 ii Declaration I declare that that the work contained within this thesis is an account of my own research, except where work by others published or unpublished is noted, while I was enrolled in the Bachelor of Science with Honours degree at Murdoch University, Western Australia. This work has not been previously submitted for a degree at any institution. Stephen Seaton November 2012 iii Conference Presentations Seaton, S.A.H., Matusick, G., Hardy, G. 2012. Drought induced tree collapse and the outbreak of Phoracantha semipunctata poses a risk for forest under climate change. Abstract presented at the Combined Biological Sciences Meeting (CBSM) 2012, 24th of August. University Club, University of Western Australia. Seaton, S.A.H., Matusick, G., Hardy, G. 2012. Occurrence of Eucalyptus longicorn borer (Phoracantha semipunctata) in the Northern Jarrah Forest following severe drought. To be presented at The Australian Entomological Society - 43rd AGM & Scientific Conference and Australasian Arachnological Society - 2012 Conference. 25th – 28th November. The Old Woolstore, Hobart. iv Acknowledgments I greatly appreciate the guidance, enthusiasm and encouragement and tireless support from my supervisors Dr George Matusick and Prof Giles Hardy in the Centre of Excellence for Climate Change Forests and Woodland Health. I particularly appreciate the interaction and productive discussions regarding forest ecology and entomology and proof reading the manuscript. -

Key to the Genera of Cerambycidae of Western North America
KEY TO THE GENERA OF THE CERAMBYCIDAE OF WESTERN NORTH AMERICA Version 030120 JAMES R. LaBONTE JOSHUA B. DUNLAP DANIEL R. CLARK THOMAS E. VALENTE JOSHUA J. VLACH OREGON DEPARTMENT OF AGRICULTURE Begin key Contributions and Acknowledgements James R. LaBonte, ODA (Oregon Department of Agriculture: Design and compilation of this identification aid. Joshua B. Dunlap: Acquisition of most of the images. Daniel R. Clark: Design input and testing. Thomas E. Valente, ODA: Design input and testing. Joshua J. Vlach, ODA: Design input and testing. Thomas Shahan, Thomas Valente, Steve Valley – additional images ODA: Use of the imaging system, the entomology museum, and general support. Our appreciation to USDA Forest Service and ODA for funding this project. Introduction Begin key This identification aid is a comprehensive key to the genera of western North American Cerambycidae (roundheaded or long- horned wood borers). It also includes several genera (and species) that are either established in the region or that are targets of USDA and other exotic cerambycid surveys. Keys to commonly trapped or encountered (based on ODA’s years of wood borer surveys) indigenous species are also included. *This aid will be most reliable west of the Rocky Mountains. It may not function well with taxa found in the desert West and east of the Rockies. This aid is designed to be used by individuals with a wide range of taxonomic expertise. Images of all character states are provided. Begin key Use of This Key: I This key is designed like a traditional dichotomous key, with couplets. However, PowerPoint navigational features have been used for efficiency. -

31 First Record of Batocera Rufomaculata (De Geer, 1775) from Sunderban Biosphere Reserve, West Bengal
International Journal of Entomology Research ISSN: 2455-4758 www.entomologyjournals.com Volume 1; Issue 3; March 2016; Page No. 31-32 First record of Batocera rufomaculata (De Geer, 1775) from Sunderban biosphere reserve, West Bengal 1 Bulganin Mitra, 2 Udipta Chakraborti, 3 Olive Biswas, 4 Sankarsan Roy, 5 Kaushik Mallick, 6 Priyanka Das 1, 2, 3, 4, 6 Zoological Survey of India, Prani Vigyan Bhawan, M-Block, New Alipore, Kolkata. 5 Post Graduate Department of Zoology, Asutosh College, Kolkata Abstract Studies on Longhorn beetles (Coleoptera) in Sunderban region is very poor. Altogether, 8 species under 3 subfamilies are already reported from Sunderban Biosphere Reserve. Present communication reports Batocera rufomaculata (De Geer, 1775) for the first time from this Biosphere reserve. Keywords: Sunderban Biosphere Reserve, Cerambycidae, Lamiinae, Batocera Introduction Sunderban region in India is 9600 sq km (4200 sq km of Reserved Forest and 5400 sq km of non-forest, inhabited region) which constitutes the Sunderban Biosphere Reserve (SBR). Indian Sunderban is bound on the west by river Muriganga and on the east by rivers Harinbhahga and Raimangal. Administrative boundary of the Sunderban is spread over two districts i.e. North 24-Parganas (Hingalganj, Hasnabad, Haroa, Sandeskhali - I,II, and Minakhan blocks) and South 24-Parganas (Sagar, Namkhana, Kakdwip, Patharpratima, Kultali, Mathurapur-I,II, Jaynagar-I,II, Canning-I,II, Basanti and Gosaba blocks).The extent of mangrove Reserve Forests in Indian Sunderban is around 4260 sq km, out of which 55% is under land vegetation cover and balance 45% is under water body/ inter-tidal zone. Studies on beetles and weevils (Coleoptera) in Sunderban region is very poor. -

4 Reproductive Biology of Cerambycids
4 Reproductive Biology of Cerambycids Lawrence M. Hanks University of Illinois at Urbana-Champaign Urbana, Illinois Qiao Wang Massey University Palmerston North, New Zealand CONTENTS 4.1 Introduction .................................................................................................................................. 133 4.2 Phenology of Adults ..................................................................................................................... 134 4.3 Diet of Adults ............................................................................................................................... 138 4.4 Location of Host Plants and Mates .............................................................................................. 138 4.5 Recognition of Mates ................................................................................................................... 140 4.6 Copulation .................................................................................................................................... 141 4.7 Larval Host Plants, Oviposition Behavior, and Larval Development .......................................... 142 4.8 Mating Strategy ............................................................................................................................ 144 4.9 Conclusion .................................................................................................................................... 148 Acknowledgments ................................................................................................................................. -

Pest and Diseases in Mango (Mangifera Indica L.) J
PEST AND DISEASES IN MANGO (MANGIFERA INDICA L.) J. González-Fernández, J.I. Hormaza IHSM la Mayora CSIC-UMA, 29750 Algarrobo, Malaga, Spain EXECUTIVE SUMMARY In this work, we review the most important pests and diseases that affect mango production worldwide as well as the main measures implemented to control them. Pests and diseases are the main factors that can impact sustainable mango fruit production in the tropics and subtropics worldwide. Commercial cultivation of mango, characterized by expansion to new areas, changing crop management, replacement of varieties and increased chemical interventions, has altered significantly the pest and disease community structure in this crop in the different mango producing regions. In addition, climate change is inducing the emergence of new pests and, whereas globalization and trade liberalization have created wide opportunities for mango commercialization growth, at the same time, this can result in faster dispersion of pests and diseases among different mango growing areas if proper sanitary measures are not implemented. This review covers different topics related to pests and diseases in mango. First, a thorough description of the main pests and diseases that affect mango is provided. Second, the different approaches used in different mango producing countries for chemical and biological control are described. Third, recommendations for appropriate mango management techiques that include integrated pest and disease management, reduction in the use of chemicals and the implementation of a good monitoring and surveillance system to help control the main pests and diseases, are also discussed. Finally, the current knowledge on agrohomeopathy and Korean Natural Farming is analyzed and recommendations on future lines of research to optimize mango pest and disease control are discussed. -

Transcriptome and Gene Expression Analysis of Rhynchophorus Ferrugineus (Coleoptera: Curculionidae) During Developmental Stages
Transcriptome and gene expression analysis of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) during developmental stages Hongjun Yang1,2, Danping Xu1, Zhihang Zhuo1,2,3, Jiameng Hu2 and Baoqian Lu4 1 College of Life Science, China West Normal University, Nanchong, Sichuan, China 2 Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Key Laboratory of Germplasm Resources Biology of Tropical Special Ornamental Plants of Hainan Province, College of Forestry, Hainan University, Haikou, Hainan, China 3 Key Laboratory of Integrated Pest Management on Crops in South China, Ministry of Agriculture, South China Agricultural University, Guangzhou, Guangdong, China 4 Key Laboratory of Integrated Pest Management on Tropical Crops, Ministry of Agriculture China, Environ- ment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China ABSTRACT Background. Red palm weevil, Rhynchophorus ferrugineus Olivier, is one of the most destructive pests harming palm trees. However, genomic resources for R. ferrugineus are still lacking, limiting the ability to discover molecular and genetic means of pest control. Methods. In this study, PacBio Iso-Seq and Illumina RNA-seq were used to generate transcriptome from three developmental stages of R. ferrugineus (pupa, 7th-instar larva, adult) to increase the understanding of the life cycle and molecular characteristics of the pest. Results. Sequencing generated 625,983,256 clean reads, from which 63,801 full-length transcripts were assembled with N50 of 3,547 bp. Expression analyses revealed 8,583 differentially expressed genes (DEGs). Moreover, gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis revealed that these Submitted 5 March 2020 Accepted 29 September 2020 DEGs were mainly related to the peroxisome pathway which associated with metabolic Published 2 November 2020 pathways, material transportation and organ tissue formation. -

Studies on Wood Boring Insects of Malabar Region of Kerala
Vol. 5, Issue 1, Pp: (19-25), March, 2021 STUDIES ON WOOD BORING INSECTS OF MALABAR REGION OF KERALA GANA KARIKKAN1 AND SHEIK MOHAMMED SHAMSUDEEN2 1Forest Entomology and GIS Research Laboratory, Department of Zoology, Sir Syed College, Taliparamba, Kannur, Kerala- 670142, India 2Department of Zoology, Mananthavady Campus, Kannur University, Wayanad, Kerala- 670645, India ARTICLE INFORMATION ABSTRACT Article History: The present study of wood boring insect was carried out during the period th Received : 20 March 2021 from February 2019 to January 2020. A Field survey was conducted in Accepted: 18th April 2021 Published online: 17th May 2021 different areas -Thaliparamba, Kannadiparamba, Thottada, Aralam Wildlife Sanctuary (Pookkund, Thullal, Valayamchal) and Elayavoor of Kannur Author’s contribution District. This short-term study recorded 38 species of wood borers. The GK complied all the results and performed the experiments, SMS collected specimens were under 7 Families named Cerambycidae, designed the study and wrote the paper. Curculionidae, Brentidae, Bostrychidae, Platypodidae, Buprestidae and Lyctidae. Cerambycidae was the dominant family followed by Bostrychidae Key words: and least was Brentidae. The collected specimens were pinned, dried and Wood boring insect, survey, Thaliparamba, Wildlife Sanctuary, stored in insect cabinets and kept in Forest Entomology and GIS Research Cerambycidae, Curculionidae, Brentidae Laboratory. 2. MATERIALS AND METHODS 1. INTRODUCTION In Kerala about 53 species of beetles were recorded The present study of wood boring insect was carried as pest of one or more of 46 species of stored timber out during the period from February 2019 to January (Mathew, 1982). Studies on the timber beetles found 2020. Different locations in Malabar region were in the Indian subcontinent were primarily made by selected for the investigation. -

Ethnoentomological and Distributional Notes on Cerambycidae and Other Coleoptera of Guerrero and Puebla,Mexico
The Coleopterists Bulletin, 71(2): 301–314. 2017. ETHNOENTOMOLOGICAL AND DISTRIBUTIONAL NOTES ON CERAMBYCIDAE AND OTHER COLEOPTERA OF GUERRERO AND PUEBLA,MEXICO JONATHAN D. AMITH Research Affiliate, Department of Anthropology, Gettysburg College, Campus Box 2895, Gettysburg, PA 17325, U.S.A. and Research Associate, National Museum of Natural History, Smithsonian Institution, Washington, DC 20013-7012, U.S.A. AND STEVEN W. LINGAFELTER Systematic Entomology Laboratory, Agriculture Research Service, United States Department of Agriculture, National Museum of Natural History, Smithsonian Institution,Washington, DC 20013-7012, U.S.A. Current address: 8920 South Bryerly Ct., Hereford, AZ 85615, U.S.A. ABSTRACT This article presents both ethnoentomological notes on Nahuatl and Mixtec language terms as they are applied to Cerambycidae (Coleoptera) and distributional records for species collected during three projects carried out in the states of Guerrero and Puebla, Mexico. Some comparative data from other Mesoamerican and Native American languages are discussed. Indigenous common names are mapped onto current taxonomic nomenclature, and an analysis is offered of the logical basis for Indigenous classification: the exclusion of some cerambycids and the inclusion of other beetles in the nominal native “cerambycid” category. New state distributional records for the Cerambycidae collected in this study are offered for Guerrero: Bebelis picta Pascoe, Callipogon senex Dupont, Neocompsa macrotricha Martins, Olenosus ser- rimanus Bates, Ornithia mexicana zapotensis Tippmann, Stenygra histrio Audinet-Serville, Strongylaspis championi Bates, Lissonotus flavocinctus puncticollis Bates, and Nothopleurus lobigenis Bates; and Puebla: Juiaparus mexicanus (Thomson), Ptychodes guttulatus Dillon and Dillon, and Steirastoma senex White. Key Words: linguistics, etymology, Nahuatl, Mixtec, longhorned beetle, wood-borer DOI.org/10.1649/0010-065X-71.2.301 The present article emerges from two language shapes. -

Chapter 11. Host Preference Testing for Parasitoids of a Eucalyptus Borer in California
ASSESSING HOST RANGES OF PARASITOIDS AND PREDATORS _________________________________ CHAPTER 11. HOST PREFERENCE TESTING FOR PARASITOIDS OF A EUCALYPTUS BORER IN CALIFORNIA T. D. Paine,1 J. G. Millar,1 and L. M. Hanks2 1Department of Entomology, University of California, Riverside, California [email protected] 2Department of Entomology, University of Illinois, Urbana, Illinois DESCRIPTION OF PEST INVASION AND PROBLEM Of the more than 700 species in the genus Eucalyptus L’Heritier native to Australia and New Guinea, approximately 90 species have been introduced into North America over the last 150 years (Doughty, 2000). Eucalyptus trees were first propagated in California from seed brought from Australia. Insect pests and diseases associated with living trees were not introduced with the seeds. As a result, the trees growing in California were relatively free of pests until the last two decades of the twentieth century (Paine and Millar, 2002). Phoracantha semipunctata (Fabricius) (Coleoptera: Cerambycidae) is native to Australia but has been accidentally introduced into virtually all of the Eucalyptus-growing regions of the world, including California, and is causing significant tree mortality in many of those areas (Paine et al., 1993, 1995, 1997). The beetles are attracted to volatile chemical cues produced by downed Eucalyptus and Angophora Cav. trees, broken branches, or standing stressed trees that are suitable larval host material (Chararas, 1969; Drinkwater, 1975; Ivory, 1977; Gonzalez- Tirado, 1987; Hanks et al., 1991). After mating on the bark surface, females oviposit under loose, exfoliated bark. The neonate larvae mine through the outer bark and feed in the nutri- tious inner bark, cambium, and outer layers of xylem (Hanks et al., 1993). -

Catalogue of Afghanistan Longhorn Beetles (Coleoptera, Cerambycidae) with Two Descriptions of New Phytoecia (Parobereina Danilevsky, 2018) from Central Asia
Humanity space International almanac VOL. 8, No 2, 2019: 104-140 http://zoobank.org/urn:lsid:zoobank.org:pub:30F6FA0A-2D7A-4ED2-9EAE-AB7707FFBE61 Catalogue of Afghanistan Longhorn beetles (Coleoptera, Cerambycidae) with two descriptions of new Phytoecia (Parobereina Danilevsky, 2018) from Central Asia M.A. Lazarev State Budget Professional Educational Institution of the Moscow Region “Chekhov technical college” Novaya str., 4, Novyi Byt village, Chekhov District, Moscow Region 142322 Russia e-mail: [email protected]; [email protected] Key words: Coleoptera, Cerambycidae, taxonomy, distribution, new species, Afghanistan, Pakistan. Abstract: The Catalogue includes all 78 Cerambycidae species of Afghanistan fauna known up to 2019 with the references to the original descriptions; 22 species were not mentioned for Afghanistan in Palaearctic Cerambycidae Catalogue by Löbl & Smetana (2010). Bibliography of each species usually includes the geographical information from corresponding publications. Many new taxonomy positions published after 2010 are used here without special remarks. Agapanthia (Epoptes) dahli ustinovi Danilevsky, 2013 stat. nov. is downgraded from the species level. Two species are described as new Phytoecia (Parobereina) pashtunica sp. n. from Afghanistan and Phytoecia (Parobereina) heinzi sp.n. from Pakistan. The present work is an attempt to summarize all data published up to now on Cerambycidae of Afghanistan fauna. Family CERAMBYCIDAE Latreille, 1802 subfamily Prioninae Latreille, 1802 tribe Macrotomini J. Thomson, 1861 genus Anomophysis Quentin & Villiers, 1981: 374 type species Prionus spinosus Fabricius, 1787 inscripta C.O. Waterhouse, 1884: 380 (Macrotoma) Heyrovský, 1936: 211 - Wama; Tippmann, 1958: 41 - Kabul, Ost- Afghanistan, 1740; Sarobi, am Kabulflus, 900 m; Mangul, Bashgultal, Nuristan, Ost-Afghanistan, 1250 m; Fuchs, 1961: 259 - Sarobi 1100 m, O.-Afghanistan; Fuchs, 1967: 432 - Afghanistan, 25 km N von Barikot, 1800 m, Nuristan; Nimla, 40 km SW von Dschelalabad; Heyrovský, 1967: 156 - Zentral-Afghanistan, Prov. -
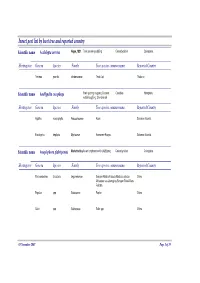
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

20 Pest Management in Organic Cacao
20 Pest Management in Organic Cacao Régis Babin* International Centre of Insect Physiology and Ecology (icipe), Nairobi, Kenya Introduction Americas produced around 14% of total world production of cocoa (FAOSTAT, 2014). General information on cacao Cacao crop expansion in Africa and Asia came with the emergence of major pests Cacao, Theobroma cacao, is a small tree and diseases, which have adapted to the from the family Malvaceae, and originated crop from their local host plants. The most in different forest areas of South and Central infamous examples are the cocoa mirids America (Wood, 1985). During the 20th cen- Sahlbergella singularis Hagl. and Distantiel- tury, the cacao-growing belt spread consid- la theobroma Dist. (Hemiptera: Miridae), and erably over tropical areas of America, Africa the black pod disease due to Phytophthora and Asia, and is around 10 million ha today palmivora Butler and Phytophthora mega- (FAOSTAT, 2014). Cocoa beans are pro- karya, which became major threats for West duced for butter and powder that are used African-producing countries in the 1960s mainly in chocolate manufacture. In 2014, and 1970s, respectively (Entwistle, 1985; chocolate confectionery produced revenues Lass, 1985). In Latin America, witches’ of around US$120 bn, and these are ex- broom disease due to the basidiomycete fun- pected to grow with the developing markets gus Moniliophthora perniciosa highly im- in countries with rising middle classes pacted production of cocoa in Brazil in the (Hawkins and Chen, 2014). At the same time, 1990s (Meinhardt et al., 2008), while the cocoa world production rose constantly for frosty pod rot, due to Moniliophthora roreri, decades and reached 5 million t in 2012 that is widely spread in Latin America, cur- (FAOSTAT, 2014).