4 Reproductive Biology of Cerambycids
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

4Th National IPM Symposium
contents Foreword . 2 Program Schedule . 4 National Roadmap for Integrated Pest Management (IPM) . 9 Whole Systems Thinking Applied to IPM . 12 Fourth National IPM Symposium . 14 Poster Abstracts . 30 Poster Author Index . 92 1 foreword Welcome to the Fourth National Integrated Pest Management The Second National IPM Symposium followed the theme “IPM Symposium, “Building Alliances for the Future of IPM.” As IPM Programs for the 21st Century: Food Safety and Environmental adoption continues to increase, challenges facing the IPM systems’ Stewardship.” The meeting explored the future of IPM and its role approach to pest management also expand. The IPM community in reducing environmental problems; ensuring a safe, healthy, has responded to new challenges by developing appropriate plentiful food supply; and promoting a sustainable agriculture. The technologies to meet the changing needs of IPM stakeholders. meeting was organized with poster sessions and workshops covering 22 topic areas that provided numerous opportunities for Organization of the Fourth National Integrated Pest Management participants to share ideas across disciplines, agencies, and Symposium was initiated at the annual meeting of the National affiliations. More than 600 people attended the Second National IPM Committee, ESCOP/ECOP Pest Management Strategies IPM Symposium. Based on written and oral comments, the Subcommittee held in Washington, DC, in September 2001. With symposium was a very useful, stimulating, and exciting experi- the 2000 goal for IPM adoption having passed, it was agreed that ence. it was again time for the IPM community, in its broadest sense, to come together to review IPM achievements and to discuss visions The Third National IPM Symposium shared two themes, “Putting for how IPM could meet research, extension, and stakeholder Customers First” and “Assessing IPM Program Impacts.” These needs. -

Asian Long-Horned Beetle Anoplophora Glabripennis
MSU’s invasive species factsheets Asian long-horned beetle Anoplophora glabripennis The Asian long-horned beetle is an exotic wood-boring insect that attacks various broadleaf trees and shrubs. The beetle has been detected in a few urban areas of the United States. In Michigan, food host plants for this insect are abundantly present in urban landscapes, hardwood forests and riparian habitats. This beetle is a concern to lumber, nursery, landscaping and tourism industries. Michigan risk maps for exotic plant pests. Other common name starry sky beetle Systematic position Insecta > Coleoptera > Cerambycidae > Anoplophora glabripennis (Motschulsky) Global distribution Native to East Asia (China and Korea). Outside the native range, the beetle infestation has been found in Austria and Canada (Toronto) and the United States: Illinois (Chicago), New Jersey, New York (Long Island), and Asian long-horned beetle. Massachusetts. Management notes Quarantine status The only effective eradication technique available in This insect is a federally quarantined organism in North America has been to cut and completely destroy the United States (NEPDN 2006). Therefore, detection infested trees (Cavey 2000). must be reported to regulatory authorities and will lead to eradication efforts. Economic and environmental significance Plant hosts to Michigan A wide range of broadleaf trees and shrubs including If the beetle establishes in Michigan, it may lead to maple (Acer spp.), poplar (Populus spp.), willow (Salix undesirable economic consequences such as restricted spp.), mulberry (Morus spp.), plum (Prunus spp.), pear movements and exports of solid wood products via (Pyrus spp.), black locust (Robinia pseudoacacia) and elms quarantine, reduced marketability of lumber, and reduced (Ulmus spp.). -

Poplars and Willows: Trees for Society and the Environment / Edited by J.G
Poplars and Willows Trees for Society and the Environment This volume is respectfully dedicated to the memory of Victor Steenackers. Vic, as he was known to his friends, was born in Weelde, Belgium, in 1928. His life was devoted to his family – his wife, Joanna, his 9 children and his 23 grandchildren. His career was devoted to the study and improve- ment of poplars, particularly through poplar breeding. As Director of the Poplar Research Institute at Geraardsbergen, Belgium, he pursued a lifelong scientific interest in poplars and encouraged others to share his passion. As a member of the Executive Committee of the International Poplar Commission for many years, and as its Chair from 1988 to 2000, he was a much-loved mentor and powerful advocate, spreading scientific knowledge of poplars and willows worldwide throughout the many member countries of the IPC. This book is in many ways part of the legacy of Vic Steenackers, many of its contributing authors having learned from his guidance and dedication. Vic Steenackers passed away at Aalst, Belgium, in August 2010, but his work is carried on by others, including mem- bers of his family. Poplars and Willows Trees for Society and the Environment Edited by J.G. Isebrands Environmental Forestry Consultants LLC, New London, Wisconsin, USA and J. Richardson Poplar Council of Canada, Ottawa, Ontario, Canada Published by The Food and Agriculture Organization of the United Nations and CABI CABI is a trading name of CAB International CABI CABI Nosworthy Way 38 Chauncey Street Wallingford Suite 1002 Oxfordshire OX10 8DE Boston, MA 02111 UK USA Tel: +44 (0)1491 832111 Tel: +1 800 552 3083 (toll free) Fax: +44 (0)1491 833508 Tel: +1 (0)617 395 4051 E-mail: [email protected] E-mail: [email protected] Website: www.cabi.org © FAO, 2014 FAO encourages the use, reproduction and dissemination of material in this information product. -

Transcriptome and Gene Expression Analysis of Rhynchophorus Ferrugineus (Coleoptera: Curculionidae) During Developmental Stages
Transcriptome and gene expression analysis of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) during developmental stages Hongjun Yang1,2, Danping Xu1, Zhihang Zhuo1,2,3, Jiameng Hu2 and Baoqian Lu4 1 College of Life Science, China West Normal University, Nanchong, Sichuan, China 2 Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Key Laboratory of Germplasm Resources Biology of Tropical Special Ornamental Plants of Hainan Province, College of Forestry, Hainan University, Haikou, Hainan, China 3 Key Laboratory of Integrated Pest Management on Crops in South China, Ministry of Agriculture, South China Agricultural University, Guangzhou, Guangdong, China 4 Key Laboratory of Integrated Pest Management on Tropical Crops, Ministry of Agriculture China, Environ- ment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China ABSTRACT Background. Red palm weevil, Rhynchophorus ferrugineus Olivier, is one of the most destructive pests harming palm trees. However, genomic resources for R. ferrugineus are still lacking, limiting the ability to discover molecular and genetic means of pest control. Methods. In this study, PacBio Iso-Seq and Illumina RNA-seq were used to generate transcriptome from three developmental stages of R. ferrugineus (pupa, 7th-instar larva, adult) to increase the understanding of the life cycle and molecular characteristics of the pest. Results. Sequencing generated 625,983,256 clean reads, from which 63,801 full-length transcripts were assembled with N50 of 3,547 bp. Expression analyses revealed 8,583 differentially expressed genes (DEGs). Moreover, gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis revealed that these Submitted 5 March 2020 Accepted 29 September 2020 DEGs were mainly related to the peroxisome pathway which associated with metabolic Published 2 November 2020 pathways, material transportation and organ tissue formation. -

Arhopalus Asperatus – a Common Longhorned Beetle
Colorado Insect of Interest Arhopalus asperatus – A Common Longhorned Beetle Scientific Name: Arhopalus asperatus (LeConte) Order: Coleoptera (Beetles) Family: Cerambycidae (Longhorned Beetles) Identification and Descriptive Features: Arhopalus asperatus is a elongate-bodied longhorned beetle that is uniformly dark gray or brown. They can be moderately large but show considerable size variation, ranging from 18-33 mm in length. The antennae are about 1/2- 3/4 the body length and are longer in males. Larvae, type of roundheaded borer, are marked by a pair of Figure 1. Arhopalus asperatus. sharp spines that curve inward found on the hind segment of the body. Distribution in Colorado: Arhopalus asperatus appears to be widely distributed throughout the forested areas of the state. Life History and Habits: Arhopalus asperatus, along with the various “pine sawyers” (Monochamous spp.), are the most common longhorned beetles associated with recently dead or felled conifers. Most conifers, including most pines, firs, douglas-fir and spruce, are known hosts. Adult lay eggs in deep bark crevices. Larvae originally feed in the cambium, later moving to sapwood and heartwood where they pack the tunnels with coarse sawdust frass. Stumps and large roots are often the most common site of larval development. Development is thought to take 2-3 years to complete. Adults are attracted to recently scorched wood following forest fires. They also will often be seen around campfires. Related Species: Other Arhopalus species are known from Colorado. Museum records for A. rusticus montanus LeConte include El Paso, Jefferson, Boulder, Mesa, and Moffat counties, suggesting widespread distribution within Colorado. It is a lighter colored species than is A. -

New Longhorn Beetles (Coleoptera: Cerambycidae) from Serbia
Arch. Biol. Sci., Belgrade, 57 (4), 27P-28P, 2005. NEW LONGHORN BEETLES (COLEOPTERA: CERAMBYCIDAE) FROM SERBIA. Nataša Pil1 and D. Stojanović2. 1Institute for Nature Conservation of Serbia, 21000 Novi Sad, Serbia and Montenegro, 2”Fruška Gora” National Park, 21208 Sremska Kamenica, Serbia and Montenegro UDC 597.76(497.11) Since the 1980’s, longhorn beetles (Coleoptera, Cerambycidae) They feed in the central region of the cone or occasionally in have been only randomly researched in Serbia. From earlier the base of old scales. The life cycle probably last two years, years, there are very detailed publications on this insect group and pupation very likely occurs in the soil. Adults emerge in (A d a m o v i ć , 1965; M i k š i ć and G e o r g i j e v i ć , 1971; April-July, on flowers. The given species differs from the simi- 1973; M i k š i ć and K o r p i č , 1985). lar Cortodera humeralis (Schaller, 1783) in having only sparse pubescence on the pronotum and head, with glabrous median The most recent data (I l i ć , 2005) indicate the presence line, and sparse pubescence on the outer border of the eye and of 245 longhorn beetle species (Coleoptera: Cerambycidae) in base of the antennae. Serbia. Not included in the mentioned publication, the follow- ing five species should be added to the list: Cortodera discolor 3. Vadonia hirsuta (Daniel and Daniel,1891) Fairmaire, 1866; Stenopterus similatus Holzschuh, 1979; Chlo- rophorus aegyptiacus (Fabricius, 1775); Agapanthia osmanlis (New data: Mt. -

Ethanol and (–)-A-Pinene: Attractant Kairomones for Some Large Wood-Boring Beetles in Southeastern USA
J Chem Ecol (2006) DOI 10.1007/s10886-006-9037-8 Ethanol and (–)-a-Pinene: Attractant Kairomones for Some Large Wood-Boring Beetles in Southeastern USA Daniel R. Miller Received: 12 September 2005 /Revised: 12 December 2005 /Accepted: 2 January 2006 # Springer Science + Business Media, Inc. 2006 Abstract Ethanol and a-pinene were tested as attractants for large wood-boring pine beetles in Alabama, Florida, Georgia, North Carolina, and South Carolina in 2002–2004. Multiple-funnel traps baited with (j)-a-pinene (released at about 2 g/d at 25–28-C) were attractive to the following Cerambycidae: Acanthocinus nodosus, A. obsoletus, Arhopalus rusticus nubilus, Asemum striatum, Monochamus titillator, Prionus pocularis, Xylotrechus integer, and X. sagittatus sagittatus. Buprestis lineata (Buprestidae), Alaus myops (Elateridae), and Hylobius pales and Pachylobius picivorus (Curculionidae) were also attracted to traps baited with (j)-a-pinene. In many locations, ethanol synergized attraction of the cerambycids Acanthocinus nodosus, A. obsoletus, Arhopalus r. nubilus, Monochamus titillator, and Xylotrechus s. sagittatus (but not Asemum striatum, Prionus pocularis,orXylotrechus integer)to traps baited with (j)-a-pinene. Similarly, attraction of Alaus myops, Hylobius pales, and Pachylobius picivorus (but not Buprestis lineata) to traps baited with (j)-a- pinene was synergized by ethanol. These results provide support for the use of traps baited with ethanol and (j)-a-pinene to detect and monitor common large wood- boring beetles from the southeastern region of the USA at ports-of-entry in other countries, as well as forested areas in the USA. Keywords Cerambycidae . Xylotrechus . Monochamus . Acanthocinus Curculionidae . Hylobius . Pachylobius . Elateridae . Alaus . Ethanol a-Pinene . -

Antennal Transcriptome Analysis and Expression Profiles of Olfactory
www.nature.com/scientificreports OPEN Antennal transcriptome analysis and expression profles of olfactory genes in Anoplophora chinensis Received: 13 June 2017 Jingzhen Wang, Ping Hu, Peng Gao, Jing Tao & Youqing Luo Accepted: 27 October 2017 Olfaction in insects is essential for host identifcation, mating and oviposition, in which olfactory Published: xx xx xxxx proteins are responsible for chemical signaling. Here, we determined the transcriptomes of male and female adult antennae of Anoplophora chinensis, the citrus longhorned beetle. Among 59,357 unigenes in the antennal assembly, we identifed 46 odorant-binding proteins, 16 chemosensory proteins (CSPs), 44 odorant receptors, 19 gustatory receptors, 23 ionotropic receptors, and 3 sensory neuron membrane proteins. Among CSPs, AchiCSP10 was predominantly expressed in antennae (compared with legs or maxillary palps), at a signifcantly higher level in males than in females, suggesting that AchiCSP10 has a role in reception of female sex pheromones. Many highly expressed genes encoding CSPs are orthologue genes of A. chinensis and Anoplophora glabripennis. Notably, AchiPBP1 and AchiPBP2 showed 100% and 96% identity with AglaPBP1 and AglaPBP2 from A. glabripennis, with similar expression profles in the two species; PBP2 was highly expressed in male antennae, whereas PBP1 was expressed in all three tissues in both males and females. These results provide a basis for further studies on the molecular chemoreception mechanisms of A. chinensis, and suggest novel targets for control of A. chinensis. Anoplophora chinensis (Forster) (Coleoptera: Cerambycidae), taxonomically regarded as a synonym of Anoplophora malasiaca, is an important worldwide quarantine pest, which is native to East Asia, but which has also been introduced to North America and Europe1,2. -

25Th U.S. Department of Agriculture Interagency Research Forum On
US Department of Agriculture Forest FHTET- 2014-01 Service December 2014 On the cover Vincent D’Amico for providing the cover artwork, “…and uphill both ways” CAUTION: PESTICIDES Pesticide Precautionary Statement This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fish or other wildlife--if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Product Disclaimer Reference herein to any specific commercial products, processes, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement, recom- mendation, or favoring by the United States government. The views and opinions of wuthors expressed herein do not necessarily reflect those of the United States government, and shall not be used for advertising or product endorsement purposes. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, sex, religion, age, disability, political beliefs, sexual orientation, or marital or family status. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at 202-720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, Room 326-W, Whitten Building, 1400 Independence Avenue, SW, Washington, D.C. -

Insects That Feed on Trees and Shrubs
INSECTS THAT FEED ON COLORADO TREES AND SHRUBS1 Whitney Cranshaw David Leatherman Boris Kondratieff Bulletin 506A TABLE OF CONTENTS DEFOLIATORS .................................................... 8 Leaf Feeding Caterpillars .............................................. 8 Cecropia Moth ................................................ 8 Polyphemus Moth ............................................. 9 Nevada Buck Moth ............................................. 9 Pandora Moth ............................................... 10 Io Moth .................................................... 10 Fall Webworm ............................................... 11 Tiger Moth ................................................. 12 American Dagger Moth ......................................... 13 Redhumped Caterpillar ......................................... 13 Achemon Sphinx ............................................. 14 Table 1. Common sphinx moths of Colorado .......................... 14 Douglas-fir Tussock Moth ....................................... 15 1. Whitney Cranshaw, Colorado State University Cooperative Extension etnomologist and associate professor, entomology; David Leatherman, entomologist, Colorado State Forest Service; Boris Kondratieff, associate professor, entomology. 8/93. ©Colorado State University Cooperative Extension. 1994. For more information, contact your county Cooperative Extension office. Issued in furtherance of Cooperative Extension work, Acts of May 8 and June 30, 1914, in cooperation with the U.S. Department of Agriculture, -

Program Book
NORTH CENTRAL BRANCH Entomological Society of America 59th Annual Meeting March 28-31, 2004 President Rob Wiedenmann The Fairmont Kansas City At the Plaza 401 Ward Parkway Kansas City, MO 64112 Contents Meeting Logistics ................................................................ 2 2003-2004 Officers and Committees, ESA-NCB .............. 4 2004 North Central Branch Award Recipients ................ 8 Program ............................................................................. 13 Sunday, March 28, 2004 Afternoon ...............................................................13 Evening ..................................................................13 Monday, March 29, 2004 Morning..................................................................14 Afternoon ...............................................................23 Evening ..................................................................42 Tuesday, March 30, 2004 Morning..................................................................43 Afternoon ...............................................................63 Evening ..................................................................67 Wednesday, March 31, 2004 Morning..................................................................68 Afternoon ...............................................................72 Author Index ..............................................................73 Taxonomic Index........................................................84 Key Word Index.........................................................88 -
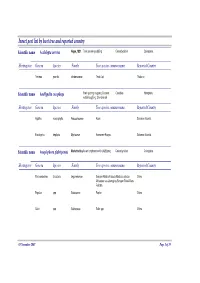
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae