NORSK ENTOMOLOGISK Tidsskriff
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Nuisance Insects and Climate Change
www.defra.gov.uk Nuisance Insects and Climate Change March 2009 Department for Environment, Food and Rural Affairs Nobel House 17 Smith Square London SW1P 3JR Tel: 020 7238 6000 Website: www.defra.gov.uk © Queen's Printer and Controller of HMSO 2007 This publication is value added. If you wish to re-use this material, please apply for a Click-Use Licence for value added material at http://www.opsi.gov.uk/click-use/value-added-licence- information/index.htm. Alternatively applications can be sent to Office of Public Sector Information, Information Policy Team, St Clements House, 2-16 Colegate, Norwich NR3 1BQ; Fax: +44 (0)1603 723000; email: [email protected] Information about this publication and further copies are available from: Local Environment Protection Defra Nobel House Area 2A 17 Smith Square London SW1P 3JR Email: [email protected] This document is also available on the Defra website and has been prepared by Centre of Ecology and Hydrology. Published by the Department for Environment, Food and Rural Affairs 2 An Investigation into the Potential for New and Existing Species of Insect with the Potential to Cause Statutory Nuisance to Occur in the UK as a Result of Current and Predicted Climate Change Roy, H.E.1, Beckmann, B.C.1, Comont, R.F.1, Hails, R.S.1, Harrington, R.2, Medlock, J.3, Purse, B.1, Shortall, C.R.2 1Centre for Ecology and Hydrology, 2Rothamsted Research, 3Health Protection Agency March 2009 3 Contents Summary 5 1.0 Background 6 1.1 Consortium to perform the work 7 1.2 Objectives 7 2.0 -
Viability of Ground Beetle Populations in Fragmented Heathlands
Viability of ground beetle populations in fragmented heathlands Henk de Vries Promotoren: Dr. L. Brussaard Hoogleraar in de Bodembiologie Dr. W. van Delden Hoogleraar in de Populatiegenetica Rijksuniversiteit Groningen Co-promotor: Dr. Th. S. van Dijk Universitair docent Biologisch Station Wijster Viability of ground beetle populations in fragmented heathlands H. H. de Vries Proefschrift ter verkrijging van de graad van doctor op gezag van de rector magnificus van de Landbouwuniversiteit Wageningen, Dr. C. M. Karssen, in het openbaar te verdedigen op woensdag 30 oktober 1996 des namiddags te vier uur in de Aula. t/)H cs"ic>zy>b LANDïîOUVV UXI VERSIT3T T ISBN 90-5485-586-X Printed on Challenger, 100% recycled paper This thesis has been accomplished at: Biological Station Wijster Wageningen Agricultural University Kampsweg 27 9418 PD Wijster The Netherlands Abstract Numbers of ground beetle species that are characteristic for heathlands were negatively associated with area, whereas this relationship was not found for the total number of ground beetle species or for unspecialised ground beetle species. In particular the number of heathland species with low dispersal ability was strongly related to area. Transplant experiments showed that some heathland species with low dispersal ability experienced reduced habitat quality in small habitats, whereas for others at least part of the unoccupied areas were of sufficient quality for successful reproduction. From the presence of occupied as well as unoccupied habitats and from knowledge on its possibilities for dispersal, it is inferred that Pterostichus lepidus lives in metapopulations with continuously and discontinuously occupied patches. Using allozymes, high levels of genetic variation were found in P. -

Fauna Lepidopterologica Volgo-Uralensis" 150 Years Later: Changes and Additions
©Ges. zur Förderung d. Erforschung von Insektenwanderungen e.V. München, download unter www.zobodat.at Atalanta (August 2000) 31 (1/2):327-367< Würzburg, ISSN 0171-0079 "Fauna lepidopterologica Volgo-Uralensis" 150 years later: changes and additions. Part 5. Noctuidae (Insecto, Lepidoptera) by Vasily V. A n ik in , Sergey A. Sachkov , Va d im V. Z o lo t u h in & A n drey V. Sv ir id o v received 24.II.2000 Summary: 630 species of the Noctuidae are listed for the modern Volgo-Ural fauna. 2 species [Mesapamea hedeni Graeser and Amphidrina amurensis Staudinger ) are noted from Europe for the first time and one more— Nycteola siculana Fuchs —from Russia. 3 species ( Catocala optata Godart , Helicoverpa obsoleta Fabricius , Pseudohadena minuta Pungeler ) are deleted from the list. Supposedly they were either erroneously determinated or incorrect noted from the region under consideration since Eversmann 's work. 289 species are recorded from the re gion in addition to Eversmann 's list. This paper is the fifth in a series of publications1 dealing with the composition of the pres ent-day fauna of noctuid-moths in the Middle Volga and the south-western Cisurals. This re gion comprises the administrative divisions of the Astrakhan, Volgograd, Saratov, Samara, Uljanovsk, Orenburg, Uralsk and Atyraus (= Gurjev) Districts, together with Tataria and Bash kiria. As was accepted in the first part of this series, only material reliably labelled, and cover ing the last 20 years was used for this study. The main collections are those of the authors: V. A n i k i n (Saratov and Volgograd Districts), S. -
Checklist of the Coleoptera of New Brunswick, Canada
A peer-reviewed open-access journal ZooKeys 573: 387–512 (2016)Checklist of the Coleoptera of New Brunswick, Canada 387 doi: 10.3897/zookeys.573.8022 CHECKLIST http://zookeys.pensoft.net Launched to accelerate biodiversity research Checklist of the Coleoptera of New Brunswick, Canada Reginald P. Webster1 1 24 Mill Stream Drive, Charters Settlement, NB, Canada E3C 1X1 Corresponding author: Reginald P. Webster ([email protected]) Academic editor: P. Bouchard | Received 3 February 2016 | Accepted 29 February 2016 | Published 24 March 2016 http://zoobank.org/34473062-17C2-4122-8109-3F4D47BB5699 Citation: Webster RP (2016) Checklist of the Coleoptera of New Brunswick, Canada. In: Webster RP, Bouchard P, Klimaszewski J (Eds) The Coleoptera of New Brunswick and Canada: providing baseline biodiversity and natural history data. ZooKeys 573: 387–512. doi: 10.3897/zookeys.573.8022 Abstract All 3,062 species of Coleoptera from 92 families known to occur in New Brunswick, Canada, are re- corded, along with their author(s) and year of publication using the most recent classification framework. Adventive and Holarctic species are indicated. There are 366 adventive species in the province, 12.0% of the total fauna. Keywords Checklist, Coleoptera, New Brunswick, Canada Introduction The first checklist of the beetles of Canada by Bousquet (1991) listed 1,365 species from the province of New Brunswick, Canada. Since that publication, many species have been added to the faunal list of the province, primarily from increased collection efforts and -

Managing for Species: Integrating the Needs of England’S Priority Species Into Habitat Management
Natural England Research Report NERR024 Managing for species: Integrating the needs of England’s priority species into habitat management. Part 2 Annexes www.naturalengland.org.uk Natural England Research Report NERR024 Managing for species: Integrating the needs of England’s priority species into habitat management. Part 2 Annexes Webb, J.R., Drewitt, A.L. and Measures, G.H. Natural England Published on 15 January 2010 The views in this report are those of the authors and do not necessarily represent those of Natural England. You may reproduce as many individual copies of this report as you like, provided such copies stipulate that copyright remains with Natural England, 1 East Parade, Sheffield, S1 2ET ISSN 1754-1956 © Copyright Natural England 2010 Project details This report results from work undertaken by the Evidence Team, Natural England. A summary of the findings covered by this report, as well as Natural England's views on this research, can be found within Natural England Research Information Note RIN024 – Managing for species: Integrating the needs of England’s priority species into habitat management. This report should be cited as: WEBB, J.R., DREWITT, A.L., & MEASURES, G.H., 2009. Managing for species: Integrating the needs of England’s priority species into habitat management. Part 2 Annexes. Natural England Research Reports, Number 024. Project manager Jon Webb Natural England Northminster House Peterborough PE1 1UA Tel: 0300 0605264 Fax: 0300 0603888 [email protected] Contractor Natural England 1 East Parade Sheffield S1 2ET Managing for species: Integrating the needs of England’s priority species into habitat i management. -
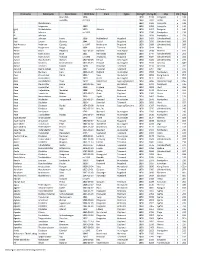
Vol5 Index.Xlsx
Vol5-Index First name Patronymic Farm Name Birth Date From Fjylke ImmigYr Immig-ID Ship Vol Page Bauerbak 1828 1850 1180 Incognito 5 108 Svendby ca 1823 1850 2412 Vesta 5 204 Abrahamsen 1850 1181 Incognito 5 108 Jorgensen 1830 1850 1252 Incognito 5 113 (girl) Mikkelsdatter 1850 Atlantic 1850 1657 Emigrant 5 146 A. Johnsen ca 1826 1850 2596 Nordpolen 5 218 A.I. Johnson 1850 2619 Nordpolen 5 219 Aad Johnsen Førre 1806 Hjelmeland Rogaland 1850 3499 (Unidentified) 5 274 Aad Larsen Østeraa 1844 Suldal Rogaland 1850 3935 (Unidentified) 5 308 Aad Andreas Pedersen Espevig ca 1847 Nedstrand Rogaland 1850 3689 (Unidentified) 5 288 Aadne Asbjørnsen Berge 1804 Kviteseid Telemark 1850 0214 Alert 5 015 Aanen Olsen Hægland 1832-03-02 Bjelland Vest-Agder 1850 0180 Hermes 5 013 Aanen Osmundsen Broe 1831 Rennesøy Rogaland 1850 3744 (Unidentified) 5 292 Aanen Osmundsen Tuestad ca 1832 Avaldsnes Rogaland 1850 3324 (Unidentified) 5 262 Aanon Abrahamsen Næsset 1817-03-28 Herad Vest-Agder 1850 3435 (Unidentified) 5 270 Aanon Knudsen Reierselmoen 1826-05-15 Froland Aust-Agder 1850 3103 Amerika 5 244+ Aanon Terjesen Kaasen 1826? Nissedal Telemark 1850 3134 Amerika 5 247 Aanund Gjermundsen Veum 1807 Fyresdal Telemark 1850 2297 Ægir 5 195 Aanund Steinsen Lien 1828-11-25 Bygland Aust-Agder 1850 2653 Nordpolen 5 222 Aasa Olavsdatter Herre 1824? Voss Hordaland 1850 0659 Kong Sverre 5 057 Aase Aslaksdatter 1810 Fjære Aust-Agder 1850 0111 Hermes 5 008 Aase Baardsdatter Thue 1827? Balestrand Sogn og Fjordane 1850 0856 Stadsraad Vogt 5 076+ Aase Clausdatter Himle 1840-02-16 -

Final Report 1
Sand pit for Biodiversity at Cep II quarry Researcher: Klára Řehounková Research group: Petr Bogusch, David Boukal, Milan Boukal, Lukáš Čížek, František Grycz, Petr Hesoun, Kamila Lencová, Anna Lepšová, Jan Máca, Pavel Marhoul, Klára Řehounková, Jiří Řehounek, Lenka Schmidtmayerová, Robert Tropek Březen – září 2012 Abstract We compared the effect of restoration status (technical reclamation, spontaneous succession, disturbed succession) on the communities of vascular plants and assemblages of arthropods in CEP II sand pit (T řebo ňsko region, SW part of the Czech Republic) to evaluate their biodiversity and conservation potential. We also studied the experimental restoration of psammophytic grasslands to compare the impact of two near-natural restoration methods (spontaneous and assisted succession) to establishment of target species. The sand pit comprises stages of 2 to 30 years since site abandonment with moisture gradient from wet to dry habitats. In all studied groups, i.e. vascular pants and arthropods, open spontaneously revegetated sites continuously disturbed by intensive recreation activities hosted the largest proportion of target and endangered species which occurred less in the more closed spontaneously revegetated sites and which were nearly absent in technically reclaimed sites. Out results provide clear evidence that the mosaics of spontaneously established forests habitats and open sand habitats are the most valuable stands from the conservation point of view. It has been documented that no expensive technical reclamations are needed to restore post-mining sites which can serve as secondary habitats for many endangered and declining species. The experimental restoration of rare and endangered plant communities seems to be efficient and promising method for a future large-scale restoration projects in abandoned sand pits. -

Notes on Some Blood-Sucking and Other Arthropods (Except Culicidae) Collected in Macedonia in 1917
153 NOTES ON SOME BLOOD-SUCKING AND OTHER ARTHROPODS (EXCEPT CULICIDAE) COLLECTED IN MACEDONIA IN 1917. By JAMES WATERSTON, Lieut. R.A.M.C., Entomologist to the Malaria Commission, Salonika. In the course of the operations for controlling mosquitos on the Macedonian front some time, necessarily limited, was devoted to collecting parasitic or noxious arthropods, and a list is here given, more particularly of those affecting man. One or two other noteworthy species are also recorded. ANOPLURA. Trichodectes subrostratus, N. Very plentiful on a kitten, Kalamaria, 8, xii. Pediculus capitis, De G., P. humanus, L., and Phthirus pubis, L. All occur. The last not commonly met with. Polyplax spinulosa, Burm. Several on a young rat, Kalamaria. RHYNCHOTA. Geocoris albipennis, F. Twice, at widely separated places, I was bitten by individuals of this species. Fortunately, on the second occasion the insect was secured without damage. Lahana, ix. Cimex lectularius, L. One of the pests of Macedonia ; only too common in wooden huts. This species drops on one's bed from roofs, unless prevented by mosquito netting. DIPTERA. Phlebotomus papatasii, Scop. Common at Kalamaria round electric lamps in huts, and also at Karasouli and Cidemli; on the Struma at Sakavca, etc. Phlebotomus perniciosus, Ann. 5 c? 14 $, Karasouli and Kalamaria, viii. Simulium sp. In hill streams near Lahana, Paprat and Oreovica larvae of one or more species of this genus were obtained, but attempts to rear the imago were unsuccessful. Pangonia marginata, F. $ (Capt. A. G. Carment). Pangonia sp. cJ, $, near variegata, F., but possibly representing two distinct species (Capt. Treadgold). Chrysops coecutiens, L. -

Lokale Energiutredninger for Kommunene I Østre Agder
Lokal energiutredning for Birkenes kommune 25/4-2012 Rolf Erlend Grundt, Agder Energi Nett Gunn Spikkeland Hansen, Rejlers Lokal energiutredning, målsetting • Forskrifter: – Forskrift om energiutredninger. (2002-12-16) – Endr. i forskrifter til energiloven. (2006-12-14) – Endr. i forskrift om energiutredninger. (2008-06- 02) • Øke kunnskapen om lokal energiforsyning, stasjonær energibruk og alternativer på dette området. • Dette for å få mer varierte energiløsninger i kommunen, og slik bidra til en samfunnsmessig rasjonell utvikling av energisystemene. • Oppdateres hvert annet år AEN ønsker også å få nytte av LEU • Oversikt over kjente utbyggingsplaner i kommunene på større tiltak til fritidsboliger, husholdninger, tjenesteyting, industri Oppdateres årlig – dette er 6. gang • Planer i kommunene der elektrisk forbruk til vann- eller romoppvarming skal erstattes av andre energibærere Gir det utslag på dimensjoneringen av nettet? Leveringspålitelighet 10 20 9 18 8 16 7 14 6 12 5 10 Birkenes 4 8 Aust-Agder 3 6 2 4 Antall perAntall rapporteringspunkt perTimer rapporteringspunkt 1 2 0 0 2007 2008 2009 2010 2007 2008 2009 2010 Gjennomsnittlig antall avbrudd Varighet på avbrudd Utførte tiltak siste to år Etablering av reserveforbindelser i Birkeland sentrum (Hauane - Valstrand). Skiftet utstyr i Birkeland TS for bedre overvåking av nettet. Kommende tiltak Fornye koblingsanlegg til Glassfiberen (3B) og etablere fjernstyring. Dette vil redusere utkoblingstid ved feil. Utføres september 2012, samarbeid med 3B. Agder Energi Nett søkte høsten 2011 om bygging av ny Vegusdal transformatorstasjon i Birkenes kommune. Gitt at konsesjon gis er stasjonen planlagt bygd imellom 2012 og 2014. Stasjonen vil blant annet legge til rette for småkraft og forbedre leveringspåliteligheten i deler av Iveland, Åmli, Froland, Birkenes og Evje- og Hornnes kommune. -

Somerset's Ecological Network
Somerset’s Ecological Network Mapping the components of the ecological network in Somerset 2015 Report This report was produced by Michele Bowe, Eleanor Higginson, Jake Chant and Michelle Osbourn of Somerset Wildlife Trust, and Larry Burrows of Somerset County Council, with the support of Dr Kevin Watts of Forest Research. The BEETLE least-cost network model used to produce Somerset’s Ecological Network was developed by Forest Research (Watts et al, 2010). GIS data and mapping was produced with the support of Somerset Environmental Records Centre and First Ecology Somerset Wildlife Trust 34 Wellington Road Taunton TA1 5AW 01823 652 400 Email: [email protected] somersetwildlife.org Front Cover: Broadleaved woodland ecological network in East Mendip Contents 1. Introduction .................................................................................................................... 1 2. Policy and Legislative Background to Ecological Networks ............................................ 3 Introduction ............................................................................................................... 3 Government White Paper on the Natural Environment .............................................. 3 National Planning Policy Framework ......................................................................... 3 The Habitats and Birds Directives ............................................................................. 4 The Conservation of Habitats and Species Regulations 2010 .................................. -

Birkenes, Herefoss Og Vegusdal PÅ DEN ANDRE SIDA
Nr. 1 2021 55. årgang BLADET MENIGHETSBirkenes, Herefoss og Vegusdal PÅ DEN ANDRE SIDA Hold ut! Husker du for et år siden, – vi trøstet oss ved å lese «Det går over» på plakater og barnetegninger. Nå blir vi minnet om «Hold ut!» Et helt år er gått, og Erna sier at vi har en bakketopp igjen! Menighetsbladet ønsker å sende varm takk til idrettslag, teplan er unnfanget i trengsel og har ingen garanti når det menigheter, foreninger, klubber og organisasjoner som gjelder gjennomføring. dette året har kjent på frustrasjon og oppgitthet. Planer og opplegg måtte til stadighet enten skrinlegges eller legges «Tilbake til normalen» er drømmen for mange. I menighe- helt om. Mye tid og krefter måtte brukes på å vurdere og tene våre er vel anledningen der til å tenke nytt, – hva skal spørre: hvordan gjør vi det, kan vi, har vi lov? Men nå altså: vi beholde og hva kan vi gjøre annerledes. Hold ut! Men uansett, – påsken skal vi beholde, og den kommer Kirka har vært lojal mot anbefalinger og forordninger, men også i år med dette grensesprengende budskapet om at «vår at det har vært lett? Særdeles nei! Men det har i alle fall ført Gud er en forsonet Gud, vår himmel er nå åpen!!!» til en del nytenking. Vi takker ansatte og frivillige i menig- God påske! hetene våre for jobben som er gjort. Vedlagte gudstjenes- RR Å, GLEDESFYLTE STUND! Å, gledesfylte stund! Var han som gav sitt liv, forblitt i dødens fengsel. Å, hvilken morgenrøde! Hva var vel da vår tro? Vår tro har sikker grunn! Men se, vår Frelser lever! Han lever! Han lever! Se livets første grøde! Han oppstod og han lever! Svein Ellingsen HOLD DEG OPPDATERT!! NESTE NUMMER AV MENIGHETSBLADET kommer ut rundt 20. -

Contribution to the Knowledge of the Fauna of Bombyces, Sphinges And
driemaandelijks tijdschrift van de VLAAMSE VERENIGING VOOR ENTOMOLOGIE Afgiftekantoor 2170 Merksem 1 ISSN 0771-5277 Periode: oktober – november – december 2002 Erkenningsnr. P209674 Redactie: Dr. J–P. Borie (Compiègne, France), Dr. L. De Bruyn (Antwerpen), T. C. Garrevoet (Antwerpen), B. Goater (Chandlers Ford, England), Dr. K. Maes (Gent), Dr. K. Martens (Brussel), H. van Oorschot (Amsterdam), D. van der Poorten (Antwerpen), W. O. De Prins (Antwerpen). Redactie-adres: W. O. De Prins, Nieuwe Donk 50, B-2100 Antwerpen (Belgium). e-mail: [email protected]. Jaargang 30, nummer 4 1 december 2002 Contribution to the knowledge of the fauna of Bombyces, Sphinges and Noctuidae of the Southern Ural Mountains, with description of a new Dichagyris (Lepidoptera: Lasiocampidae, Endromidae, Saturniidae, Sphingidae, Notodontidae, Noctuidae, Pantheidae, Lymantriidae, Nolidae, Arctiidae) Kari Nupponen & Michael Fibiger [In co-operation with Vladimir Olschwang, Timo Nupponen, Jari Junnilainen, Matti Ahola and Jari- Pekka Kaitila] Abstract. The list, comprising 624 species in the families Lasiocampidae, Endromidae, Saturniidae, Sphingidae, Notodontidae, Noctuidae, Pantheidae, Lymantriidae, Nolidae and Arctiidae from the Southern Ural Mountains is presented. The material was collected during 1996–2001 in 10 different expeditions. Dichagyris lux Fibiger & K. Nupponen sp. n. is described. 17 species are reported for the first time from Europe: Clostera albosigma (Fitch, 1855), Xylomoia retinax Mikkola, 1998, Ecbolemia misella (Püngeler, 1907), Pseudohadena stenoptera Boursin, 1970, Hadula nupponenorum Hacker & Fibiger, 2002, Saragossa uralica Hacker & Fibiger, 2002, Conisania arida (Lederer, 1855), Polia malchani (Draudt, 1934), Polia vespertilio (Draudt, 1934), Polia altaica (Lederer, 1853), Mythimna opaca (Staudinger, 1899), Chersotis stridula (Hampson, 1903), Xestia wockei (Möschler, 1862), Euxoa dsheiron Brandt, 1938, Agrotis murinoides Poole, 1989, Agrotis sp.