(Simarouba Amara and Goupia Glabra) in French Guyana Agnès Granier, R Huc, F Colin
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Towards an Understanding of the Evolution of Violaceae from an Anatomical and Morphological Perspective Saul Ernesto Hoyos University of Missouri-St
University of Missouri, St. Louis IRL @ UMSL Theses Graduate Works 8-7-2011 Towards an understanding of the evolution of Violaceae from an anatomical and morphological perspective Saul Ernesto Hoyos University of Missouri-St. Louis, [email protected] Follow this and additional works at: http://irl.umsl.edu/thesis Recommended Citation Hoyos, Saul Ernesto, "Towards an understanding of the evolution of Violaceae from an anatomical and morphological perspective" (2011). Theses. 50. http://irl.umsl.edu/thesis/50 This Thesis is brought to you for free and open access by the Graduate Works at IRL @ UMSL. It has been accepted for inclusion in Theses by an authorized administrator of IRL @ UMSL. For more information, please contact [email protected]. Saul E. Hoyos Gomez MSc. Ecology, Evolution and Systematics, University of Missouri-Saint Louis, 2011 Thesis Submitted to The Graduate School at the University of Missouri – St. Louis in partial fulfillment of the requirements for the degree Master of Science July 2011 Advisory Committee Peter Stevens, Ph.D. Chairperson Peter Jorgensen, Ph.D. Richard Keating, Ph.D. TOWARDS AN UNDERSTANDING OF THE BASAL EVOLUTION OF VIOLACEAE FROM AN ANATOMICAL AND MORPHOLOGICAL PERSPECTIVE Saul Hoyos Introduction The violet family, Violaceae, are predominantly tropical and contains 23 genera and upwards of 900 species (Feng 2005, Tukuoka 2008, Wahlert and Ballard 2010 in press). The family is monophyletic (Feng 2005, Tukuoka 2008, Wahlert & Ballard 2010 in press), even though phylogenetic relationships within Violaceae are still unclear (Feng 2005, Tukuoka 2008). The family embrace a great diversity of vegetative and floral morphologies. Members are herbs, lianas or trees, with flowers ranging from strongly spurred to unspurred. -

D:\REVISTA EIDENAR\Eidenar EDIC
CARACTERIZACIÓN ECOLÓGICA DE LA MICROCUENCA ABASTECEDORA DE AGUA DEL ACUEDUCTO MUNICIPAL DE SAN JOSÉ DEL GUAVIARE ECOLOGICAL CHARACTERIZATION OF THE MICRO-WATER SHED THAT SUPPLIES WATER TO THE ACUEDUCT SAN JOSÉ DEL GUAVIARE MONICIPALITY GUAVIARE RESUMEN Juan Carlos García-R.,Ecologo, Ph.D. Student Se examinaron la diversidad vegetal y los Massey Univerity, New Zealand macroinvertebrados acuáticos de la microcuenca La María, Departamento del Guaviare, Colombia. Fueron [email protected] establecidas dos parcelas de muestreo para determinar la diversidad y estructura vegetal del bosque protector de Luis Fernando Galindez, Ecologo. la microcuenca y cuatro estaciones para reconocer la Empo Aguas calidad del agua a través de macroinvertebrados San Jose del Giuaviare bioindicadores. Se registraron 139 individuos de 47 [email protected] especies siendo las más abundantes Socratea exorrhiza [email protected] (34 individuos), Virola calophylla (10 individuos) y Goupia glabra, Lindackeria paludosa y Oenocarpus bataua (seis individuos). Se estimó el índice biótico BMWP, para evaluar la calidad del agua, obteniendo valores aceptables para las tres primeras estaciones y entre dudosa y crítica para la última estación (posterior al ___________ embalse de distribución). Estos valores permiten deter- *Recibido: 26 Septiembre 2011 *Aceptado 25 Octubre 2011 minar un agua de buena calidad para uso potable de la pp. 31-43 Facultad de Ingeniería «EIDENAR» 31 ISSN 1692-9918 Ing. Recur. Nat. Ambient. comunidad de San José del Guaviare. gestión. Por esta razón, la definición de ecosistema es conceptualmente aplicable a los ríos, los cuales son denominados ecosistemas lóticos. PALABRAS CLAVE Los ambientes lóticos son influenciados por los ecosistemas terrestres adyacentes y por las caracterís- ticas de sus cuencas hidrográficas, tales como tipo de Macroinvertebrados acuáticos, microcuenca La María, suelo, geología, vegetación y ocupación humana. -

Evolution of Agroforestry As a Modern Science
Chapter 2 Evolution of Agroforestry as a Modern Science Jagdish C. Dagar and Vindhya P. Tewari Abstract Agroforestry is as old as agriculture itself. Many of the anecdotal agro- forestry practices, which are time tested and evolved through traditional indigenous knowledge, are still being followed in different agroecological zones. The tradi- tional knowledge and the underlying ecological principles concerning indigenous agroforestry systems around the world have been successfully used in designing the improved systems. Many of them such as improved fallows, homegardens, and park systems have evolved as modern agroforestry systems. During past four decades, agroforestry has come of age and begun to attract the attention of the international scientific community, primarily as a means for sustaining agricultural productivity in marginal lands and solving the second-generation problems such as secondary salinization due to waterlogging and contamination of water resources due to the use of excess nitrogen fertilizers and pesticides. Research efforts have shown that most of the degraded areas including saline, waterlogged, and perturbation ecolo- gies like mine spoils and coastal degraded mangrove areas can be made productive by adopting suitable agroforestry techniques involving highly remunerative compo- nents such as plantation-based farming systems, high-value medicinal and aromatic plants, livestock, fishery, poultry, forest and fruit trees, and vegetables. New con- cepts such as integrated farming systems and urban and peri-urban agroforestry have emerged. Consequently, the knowledge base of agroforestry is being expanded at a rapid pace as illustrated by the increasing number and quality of scientific pub- lications of various forms on different aspects of agroforestry. It is both a challenge and an opportunity to scientific community working in this interdisciplinary field. -

Structural Traits of an Eight Year Old Secondary Forest in the Central Amazon Preisinger, H.\ Martins, G
German-Brazilian Workshop on Neotropical Ecosystems - Achievements and Prospects of Cooperative Research Hamburg, September 3-8, 2000 Concepts and Paradigrns for Management of Ecosystem Resources - Summaries of Posters, Session 6 Structural Traits of an Eight Year Old Secondary Forest in the Central Amazon Preisinger, H.\ Martins, G. C.2 and Coêlho, L. F.3 I Universitât Hamburg, Deutschland 2 EMBRAPAAmazônia Ocidental, Manaus, Brazil 3 INPA, Manaus, Brazil In 1992, a 17 ha agro-ecological field trial was established composition of the herb layer is very distinct from the tree at the experimental site ofEMBRAPAAmazônia Ocidental, layer. The comparison of the growth- forro spectra of the situated 28 km north of Manaus, Amazonas, Brazil. The herb and the tree layer reveals that the spectra of herb and area, which was covered by an 8 year old secondary forest, tree layer are similar. Only the growth-form types with had been slashed and burned in the conventional manner. A herbaceous species ("WH" to "UH" in the graph) are absent secondary forest of 1 ha was left standing as a reference. in the tree layer or represented by only few species. The area of 100 x 100 m was divided into 10 x 10 m plots There are obviously no environmental gradients or bounda- and floristical and structural data of the vegetation of these ries in the experimental site, e.g. of soil qualities. The plots were collected, such as floristic gradients and spatial patterns must therefore be 1 Floristical composition of each of the plots, covering ali interpreted as the result of the history of the sites (= use the vascular plants (woody and herbaceous plants of ali history, mainly former disturbance events), plant ages); distributions by chance and the result of interactions and 2 Diameters of alI the trunks of each species occurring in the interspecific competition of the species. -

Chec List What Survived from the PLANAFLORO Project
Check List 10(1): 33–45, 2014 © 2014 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution What survived from the PLANAFLORO Project: PECIES S Angiosperms of Rondônia State, Brazil OF 1* 2 ISTS L Samuel1 UniCarleialversity of Konstanz, and Narcísio Department C.of Biology, Bigio M842, PLZ 78457, Konstanz, Germany. [email protected] 2 Universidade Federal de Rondônia, Campus José Ribeiro Filho, BR 364, Km 9.5, CEP 76801-059. Porto Velho, RO, Brasil. * Corresponding author. E-mail: Abstract: The Rondônia Natural Resources Management Project (PLANAFLORO) was a strategic program developed in partnership between the Brazilian Government and The World Bank in 1992, with the purpose of stimulating the sustainable development and protection of the Amazon in the state of Rondônia. More than a decade after the PLANAFORO program concluded, the aim of the present work is to recover and share the information from the long-abandoned plant collections made during the project’s ecological-economic zoning phase. Most of the material analyzed was sterile, but the fertile voucher specimens recovered are listed here. The material examined represents 378 species in 234 genera and 76 families of angiosperms. Some 8 genera, 68 species, 3 subspecies and 1 variety are new records for Rondônia State. It is our intention that this information will stimulate future studies and contribute to a better understanding and more effective conservation of the plant diversity in the southwestern Amazon of Brazil. Introduction The PLANAFLORO Project funded botanical expeditions In early 1990, Brazilian Amazon was facing remarkably in different areas of the state to inventory arboreal plants high rates of forest conversion (Laurance et al. -

Tree Height Growth in a Neotropical Rain Forest
Ecology, 82(5), 2001, pp. 1460±1472 q 2001 by the Ecological Society of America GETTING TO THE CANOPY: TREE HEIGHT GROWTH IN A NEOTROPICAL RAIN FOREST DEBORAH A. CLARK1 AND DAVID B. CLARK1 Department of Biology, University of Missouri±St. Louis, 8001 Natural Bridge Road, St. Louis, Missouri 63121-4499, USA Abstract. There is still limited understanding of the processes underlying forest dy- namics in the world's tropical rain forests, ecosystems of disproportionate importance in terms of global biogeochemistry and biodiversity. Particularly poorly documented are the nature and time scale of upward height growth during regeneration by the tree species in these communities. In this study, we assessed long-term height growth through ontogeny for a diverse group of canopy and emergent tree species in a lowland neotropical rain forest (the La Selva Biological Station, northeastern Costa Rica). Species were evaluated based on annual height measurements of large samples of individuals in all postseedling size classes, over a 16-yr period (.11 000 increments). The study species were seven nonpi- oneers (Minquartia guianensis, Lecythis ampla, Hymenolobium mesoamericanum, Sima- rouba amara, Dipteryx panamensis, Balizia elegans, and Hyeronima alchorneoides) and two pioneers (Cecropia obtusifolia and Cecropia insignis). For each species, inherent height growth capacity was estimated as the mean of the ®ve largest annual height increments (from different individuals) in each juvenile size class (from 50 cm tall to 20 cm in diameter). All species showed marked ontogenetic increases in this measure of height growth potential. At all sizes, there were highly signi®cant differences among species in height growth potential. -

How Rare Is Too Rare to Harvest? Management Challenges Posed by Timber Species Occurring at Low Densities in the Brazilian Amazon
Forest Ecology and Management 256 (2008) 1443–1457 Contents lists available at ScienceDirect Forest Ecology and Management journal homepage: www.elsevier.com/locate/foreco How rare is too rare to harvest? Management challenges posed by timber species occurring at low densities in the Brazilian Amazon Mark Schulze a,b,c,*, James Grogan b,d, R. Matthew Landis e, Edson Vidal b,f a School of Forest Resources and Conservation, University of Florida, P.O. Box 110760, Gainesville, FL 32611, USA b Instituto do Homem e Meio Ambiente da Amazoˆnia (IMAZON), R. Domingos Marreiros, no. 2020 Bairro Fa´tima, Bele´m, Para´ 66060-160, Brazil c Instituto Floresta Tropical (IFT), Caixa Postal 13077, Bele´m, Para´ 66040-970, Brazil d Yale University School of Forestry & Environmental Studies, 360 Prospect St., New Haven, CT 06511, USA e Department of Biology, Middlebury College, Middlebury, VT 05753, USA f ESALQ/Universidade de Sa˜o Paulo, Piracicaba, Sa˜o Paulo 13.418-900, Brazil ARTICLE INFO ABSTRACT Article history: Tropical forests are characterized by diverse assemblages of plant and animal species compared to Received 11 September 2007 temperate forests. Corollary to this general rule is that most tree species, whether valued for timber or Received in revised form 28 January 2008 not, occur at low densities (<1 adult tree haÀ1) or may be locally rare. In the Brazilian Amazon, many of Accepted 28 February 2008 the most highly valued timber species occur at extremely low densities yet are intensively harvested with Keywords: little regard for impacts on population structures and dynamics. These include big-leaf mahogany Conservation (Swietenia macrophylla), ipeˆ (Tabebuia serratifolia and Tabebuia impetiginosa), jatoba´ (Hymenaea courbaril), Reduced-impact logging and freijo´ cinza (Cordia goeldiana). -

Simarouba Monograph 3/1/04 Simarouba Amara, Glauca
Simarouba Monograph 3/1/04 Simarouba amara, glauca Family: Simaroubaceae Synonyms: Quassia simarouba, Zwingera amara, Picraena officinalis, Simarouba medicinalis Other Common Names: Aceituno, bitterwood, bois blanc, bois amer, bois frene, bois negresse, caixeta, cajú-rana, cedro blanco daguilla, dysentery bark, gavilan, malacacheta, marubá, marupá, negrito, palo blanco, palo amargo, paradise tree, pitomba, robleceillo, simaba. Overview Botanical Description Simarouba is indigenous to the Amazon rainforest and other tropical areas in Mexico, Cuba, Haita, Jamaica and Central America. It grows up to 20 m high and has a trunk 50 to 80 cm in diameter. It produces bright green leaves 20 to 50 cm in length, small white flowers, and small red fruit. Ethnobotanical Uses In traditional herbal medicine systems the bark, wood and leaves of simarouba have been used for their amoebicide, analgesic, anthelmintic, antibacterial, antidysenteric, antimalarial, antimicrobial, astringent, febrifuge, stomachic, sudorific, tonic and vermifuge properties. The traditional use of graviola has been recorded in herbal medicine systems in the following countries: Guiana,1 Belize,2 Brazil,3-6 Cuba,7,8 French Guyana,9 Haiti10 and Peru.11 Summary of Traditional Uses of Simarouba:12 Bark: Anemia, anorexia, bitter,diarrhea, dysentery, dyspepsia, emmenagogue, fever, hemorrhages, internal bleeding, intestinal worms, malaria, skin sores, sores, stomach and bowel disorders, tonic, wounds. Leaf: Astringent, colitis, diarrhea, digestive, dysentery, emmenogogue,intestinal worms, malaria, skin affections. Root: Diarrhea, dysentery, flatulence, intestinal worms, malaria, stomach pain, tonic. Primary Uses Internal A bark tea is primarily used as the first line of defense for amebic dysentery and diarrhea. It’s also used for viruses.13 External The bark has been traditionally used in herbal medicine systems externally for wounds and skin sores.2,7 © Copyrighted 2004. -

Woody-Tissue Respiration for Simarouba Amara and Minquartia Guianensis, Two Tropical Wet Forest Trees with Different Growth Habits
Oecologia (1994) 100:213-220 9Springer-Verlag 1994 Michael G. Ryan 9Robert M. Hubbard Deborah A. Clark 9Robert L. Sanford, Jr. Woody-tissue respiration for Simarouba amara and Minquartia guianensis, two tropical wet forest trees with different growth habits Received: 9 February 1994 / Accepted: 26 July 1994 Abstract We measured CO 2 etflux from stems of two maintenance respiration was 54% of the total CO2 efflux tropical wet forest trees, both found in the canopy, but for Simarouba and 82% for Minquartia. For our sam- with very different growth habits. The species were ple, sapwood volume averaged 23% of stem volume Simarouba amara, a fast-growing species associated when weighted by tree size, or 40% with no size weight- with gaps in old-growth forest and abundant in sec- ing. Using these fractions, and a published estimate of ondary forest, and Minquartia guianensis, a slow-grow- aboveground dry-matter production, we estimate the ing species tolerant of low-light conditions in annual cost of woody tissue respiration for primary old-growth forest. Per unit of bole surface, CO2 efflux forest at La Selva to be 220 or 350 gCm-2year -1, averaged 1.24 gmol m -2 s-1 for Simarouba and depending on the assumed sapwood volume. These 0.83gmolm-2s -1 for Minquartia. C02 elttux was costs are estimated to be less than 13% of the gross highly correlated with annual wood production (r 2= production for the forest. 0.65), but only weakly correlated with stem diameter (r 2 = 0.22). We also partitioned the CO2 efttux into the Key words Woody tissue respiration" Maintenance functional components of construction and mainte- respiration 9Tropical wet forest trees 9Carbon balance nance respiration. -
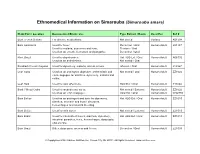
Ethnomedical Information on Simarouba (Simarouba Amara)
Ethnomedical Information on Simarouba (Simarouba amara) Plant Part / Location Documented Ethnic Use Type Extract / Route Used For Ref # Bark French Guiana For diverse medications. Not stated Various K01504 Bark Amazonia Used for fever. Decoction / Oral Human Adult L04137 Used for malaria, dysentery and tonic. Tincture / Oral Used as an emetic, hemostat, and purgative. Decoction / Oral Root Brazil Used to stop diarrhea. Hot H2O Ext / Oral Human Adult A06732 Used as an anthelmintic. Not stated / Oral Rootbark French Guyana Used for dysentery, malaria, and as a tonic. Infusion / Oral Human Adult J12967 Leaf Cuba Used as an astringent, digestive, anthelmintic and Not stated / Oral Human Adult ZZ1022 emmenogogue for diarrhea, dysentery, malaria and colitis. Leaf Haiti Used for skin affections. H2O Ext / Oral Human Adult T13846 Bark / Wood Cuba Used for wounds and sores. Not stated / External Human Adult ZZ1022 Used as an emmenagogue. H2O Ext / Oral Human Adult W02855 Bark Belize Used as an astringent and tonic for dysentery, Hot H2O Ext / Oral Human Adult ZZ1019 diarrhea, stomach and bowel disorders, hemorrhages and internal bleeding. Bark Belize Used for skin sores. Not stated / External Human Adult ZZ1019 Bark Brazil Used for intermittent fevers, diarrhea, dysentery, Hot H2O Ext / Oral Human Adult ZZ1013 intestinal parasites, tonic, hemorrhages, dyspepsia, and anemia. Bark Brazil Bitter, dyspepsia, anemia and fevers. Decoction / Oral Human Adult ZZ1099 © Copyrighted 2004. Raintree Nutrition, Inc. Carson City, NV 89701. All Rights Reserved. www.rain-tree.com Plant Part / Location Documented Ethnic Use Type Extract / Route Used For Ref # Bark + Root Brazil Used for bleeding diarrhea. Decoction / Oral Human Adult ZZ1099 Bark Brazil Used for acute and chronic dysentery, bitter tonic, Infusion / Oral Human Adult ZZ1007 febrifuge, diarrhea, inappetite, and dyspepsia. -

Amazon Plant List
Amazon Plant List The Plant list below is contributed by Dr.Christopher Dick, PhD who has worked in Amazonia for many years. Note that it is a working list and neither exhaustive nor complete. English Common Portuguese Common Plant Family Name Botanical Name Name Name Annonaceae Guatteria Envira-bobô recurvisepala Unonopsis guatterioides Myristicaceae Virola calophylla Wild nutmeg Ucuuba Iryanthera uleii Dead-bark Osteophloeum Ucuuba-amarela platyspermum Lauraceae Mezilaurus itauba Itaúba Persea americana Avocado Abacate Aniba canella Casca preciosa Aniba roseadora Pau rosa Ocotea rubra Louro-gamela Peperomia Piperaceae Ant-garden macrostachya Nymphaeaceae Victoria amazonica Amazon-lily Victoria-regia Menispermaceae Ulmaceae Trema micrantha Trema, Periquitinho Moraceae Clarisia racemosa Guariúba Naucleopsis Miratinga, Pau pica caloneura Brosimim Amapá parinarioides Cecropia Cecropiaceae Purple cecropia Imbaúba roxa purpurascens Cecropia sciadophylla Cecropia Imbaúba-torém Caruru-bravo, Bredo- Phytolaccaceae Phytolacca rivinoides Pokeweed roxo Epiphyllum Cactaceae Cactus phyllanthus Polygonaceae Coccoloba spp. Water-grape? Symeria paniculata Carauaçuzeiro Tetracera Dilleniaceae Water-vine Cipó d'agua willdenowiana Pinzona coriaceae Fire-vine Cipó-de-fôgo Caryocaraceae Caryocar villosum Piquiá Caryocar glabrum Piquiarana Margraviaceae Marcgravia Quiinaceae Clusiaceae Vismia cayennensis Lacre-branco Vismia guianensis Lacre-vermelho Symphonia Ananí used for cerol? globulifera Elaeocarpaceae Sterculiaceae Sterculia frondosa Tacacá Waltheria -

Physical Damage on Tropical Tree Saplings
Physical damage on tropical tree saplings: quantification and consequences for competition through height growth in a neotropical rain forest of French Guiana Gilles Koestel, Judy M. Rankin-de Mérona To cite this version: Gilles Koestel, Judy M. Rankin-de Mérona. Physical damage on tropical tree saplings: quantification and consequences for competition through height growth in a neotropical rain forest of French Guiana. Annales des sciences forestières, INRA/EDP Sciences, 1998, 55 (6), pp.727-742. hal-00883232 HAL Id: hal-00883232 https://hal.archives-ouvertes.fr/hal-00883232 Submitted on 1 Jan 1998 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Original article Physical damage on tropical tree saplings: quantification and consequences for competition through height growth in a neotropical rain forest of French Guiana Gilles Koestela Judy M. Rankin-de Méronab Station de recherches forestières, Inra, Centre Régional de Guyane, BP 709, 97387 Kourou cedex, France (Received 29 July 1996; revised 2 April 1997; accepted 25 May 1998) Abstract - This paper deals with the quantification and the effects of physical damage on tree regeneration dynamics in the tropical rain forest. We define physical damage as breakage resul- ting in a greater than 20 % reduction in stem diameter and its associated effects.