How Rare Is Too Rare to Harvest? Management Challenges Posed by Timber Species Occurring at Low Densities in the Brazilian Amazon
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Flexible Mating System in a Logged Population of Swietenia Macrophylla King (Meliaceae): Implications for the Management of a Threatened Neotropical Tree Species
Plant Ecol (2007) 192:169–179 DOI 10.1007/s11258-007-9322-9 ORIGINAL PAPER Flexible mating system in a logged population of Swietenia macrophylla King (Meliaceae): implications for the management of a threatened neotropical tree species Maristerra R. Lemes Æ Dario Grattapaglia Æ James Grogan Æ John Proctor Æ Roge´rio Gribel Received: 21 February 2007 / Accepted: 22 May 2007 / Published online: 19 June 2007 Ó Springer Science+Business Media B.V. 2007 Abstract Microsatellites were used to evaluate the crossed matings and that the remaining 6.75% had mating system of the remaining trees in a logged genotypes consistent with self-fertilisation. Apomixis population of Swietenia macrophylla, a highly valu- could be ruled out, since none of the 400 seedlings able and threatened hardwood species, in the Brazil- analysed had a multi-locus genotype identical to ian Amazon. A total of 25 open pollinated progeny its mother tree. The high estimate of the multi-locus arrays of 16 individuals, with their mother trees, were outcrossing rate (tm = 0.938 ± 0.009) using the mixed genotyped using eight highly polymorphic microsat- mating model also indicated that the population in ellite loci. Genotypic data analysis from the progeny this remnant stand of S. macrophylla was predomi- arrays showed that 373 out of the 400 seedlings nantly allogamous. The relatively large difference (93.25%) were unambiguously the result of out- between the multi-locus and single-locus outcrossing estimates (tmÀts = 0.117 ± 0.011) provides evidence that, in spite of the high outcrossing rate, a consid- & M. R. Lemes ( ) Á R. -

Estimating Canopy Structure in an Amazon Forest from Laser Range Finder and IKONOS Satellite Observations1
BIOTROPICA 34(4): 483±492 2002 Estimating Canopy Structure in an Amazon Forest from Laser Range Finder and IKONOS Satellite Observations1 Gregory P. Asner 2 Department of Global Ecology, Carnegie Institution of Washington, Stanford University, Stanford, California 94305, U.S.A. Michael Palace Complex Systems Research Center, Morse Hall, University of New Hampshire, Durham, New Hampshire 03824, U.S.A. Michael Keller Complex Systems Research Center, Morse Hall, University of New Hampshire, Durham, New Hampshire 03824, U.S.A. and USDA Forest Service, International Institute of Tropical Forestry, Rio Piedras, Puerto Rico, U.S.A. Rodrigo Pereira Jr. FundacËaÄo Floresta Tropical, Trv. 14 Abril, Bairro SaÄo Braz, BeleÂm CEP. 66063-140, ParaÂ, Brazil Jose N. M. Silva EMBRAPA±Amazonia Oriental, Trv. Dr Eneas Pinheiro SN, BeleÂm CEP. 66095-100, ParaÂ, Brazil and Johan C. Zweede FundacËaÄo Floresta Tropical, Trv. 14 Abril, Bairro SaÄo Braz, BeleÂm CEP. 66063-140, ParaÂ, Brazil ABSTRACT Canopy structural data can be used for biomass estimation and studies of carbon cycling, disturbance, energy balance, and hydrological processes in tropical forest ecosystems. Scarce information on canopy dimensions re¯ects the dif®- culties associated with measuring crown height, width, depth, and area in tall, humid tropical forests. New ®eld and spaceborne observations provide an opportunity to acquire these measurements, but the accuracy and reliability of the methods are unknown. We used a handheld laser range ®nder to estimate tree crown height, diameter, and depth in a lowland tropical forest in the eastern Amazon, Brazil, for a sampling of 300 trees strati®ed by diameter at breast height (DBH). -

Evolution of Agroforestry As a Modern Science
Chapter 2 Evolution of Agroforestry as a Modern Science Jagdish C. Dagar and Vindhya P. Tewari Abstract Agroforestry is as old as agriculture itself. Many of the anecdotal agro- forestry practices, which are time tested and evolved through traditional indigenous knowledge, are still being followed in different agroecological zones. The tradi- tional knowledge and the underlying ecological principles concerning indigenous agroforestry systems around the world have been successfully used in designing the improved systems. Many of them such as improved fallows, homegardens, and park systems have evolved as modern agroforestry systems. During past four decades, agroforestry has come of age and begun to attract the attention of the international scientific community, primarily as a means for sustaining agricultural productivity in marginal lands and solving the second-generation problems such as secondary salinization due to waterlogging and contamination of water resources due to the use of excess nitrogen fertilizers and pesticides. Research efforts have shown that most of the degraded areas including saline, waterlogged, and perturbation ecolo- gies like mine spoils and coastal degraded mangrove areas can be made productive by adopting suitable agroforestry techniques involving highly remunerative compo- nents such as plantation-based farming systems, high-value medicinal and aromatic plants, livestock, fishery, poultry, forest and fruit trees, and vegetables. New con- cepts such as integrated farming systems and urban and peri-urban agroforestry have emerged. Consequently, the knowledge base of agroforestry is being expanded at a rapid pace as illustrated by the increasing number and quality of scientific pub- lications of various forms on different aspects of agroforestry. It is both a challenge and an opportunity to scientific community working in this interdisciplinary field. -

Chec List What Survived from the PLANAFLORO Project
Check List 10(1): 33–45, 2014 © 2014 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution What survived from the PLANAFLORO Project: PECIES S Angiosperms of Rondônia State, Brazil OF 1* 2 ISTS L Samuel1 UniCarleialversity of Konstanz, and Narcísio Department C.of Biology, Bigio M842, PLZ 78457, Konstanz, Germany. [email protected] 2 Universidade Federal de Rondônia, Campus José Ribeiro Filho, BR 364, Km 9.5, CEP 76801-059. Porto Velho, RO, Brasil. * Corresponding author. E-mail: Abstract: The Rondônia Natural Resources Management Project (PLANAFLORO) was a strategic program developed in partnership between the Brazilian Government and The World Bank in 1992, with the purpose of stimulating the sustainable development and protection of the Amazon in the state of Rondônia. More than a decade after the PLANAFORO program concluded, the aim of the present work is to recover and share the information from the long-abandoned plant collections made during the project’s ecological-economic zoning phase. Most of the material analyzed was sterile, but the fertile voucher specimens recovered are listed here. The material examined represents 378 species in 234 genera and 76 families of angiosperms. Some 8 genera, 68 species, 3 subspecies and 1 variety are new records for Rondônia State. It is our intention that this information will stimulate future studies and contribute to a better understanding and more effective conservation of the plant diversity in the southwestern Amazon of Brazil. Introduction The PLANAFLORO Project funded botanical expeditions In early 1990, Brazilian Amazon was facing remarkably in different areas of the state to inventory arboreal plants high rates of forest conversion (Laurance et al. -

Tree Height Growth in a Neotropical Rain Forest
Ecology, 82(5), 2001, pp. 1460±1472 q 2001 by the Ecological Society of America GETTING TO THE CANOPY: TREE HEIGHT GROWTH IN A NEOTROPICAL RAIN FOREST DEBORAH A. CLARK1 AND DAVID B. CLARK1 Department of Biology, University of Missouri±St. Louis, 8001 Natural Bridge Road, St. Louis, Missouri 63121-4499, USA Abstract. There is still limited understanding of the processes underlying forest dy- namics in the world's tropical rain forests, ecosystems of disproportionate importance in terms of global biogeochemistry and biodiversity. Particularly poorly documented are the nature and time scale of upward height growth during regeneration by the tree species in these communities. In this study, we assessed long-term height growth through ontogeny for a diverse group of canopy and emergent tree species in a lowland neotropical rain forest (the La Selva Biological Station, northeastern Costa Rica). Species were evaluated based on annual height measurements of large samples of individuals in all postseedling size classes, over a 16-yr period (.11 000 increments). The study species were seven nonpi- oneers (Minquartia guianensis, Lecythis ampla, Hymenolobium mesoamericanum, Sima- rouba amara, Dipteryx panamensis, Balizia elegans, and Hyeronima alchorneoides) and two pioneers (Cecropia obtusifolia and Cecropia insignis). For each species, inherent height growth capacity was estimated as the mean of the ®ve largest annual height increments (from different individuals) in each juvenile size class (from 50 cm tall to 20 cm in diameter). All species showed marked ontogenetic increases in this measure of height growth potential. At all sizes, there were highly signi®cant differences among species in height growth potential. -

Kew Science Publications for the Academic Year 2017–18
KEW SCIENCE PUBLICATIONS FOR THE ACADEMIC YEAR 2017–18 FOR THE ACADEMIC Kew Science Publications kew.org For the academic year 2017–18 ¥ Z i 9E ' ' . -,i,c-"'.'f'l] Foreword Kew’s mission is to be a global resource in We present these publications under the four plant and fungal knowledge. Kew currently has key questions set out in Kew’s Science Strategy over 300 scientists undertaking collection- 2015–2020: based research and collaborating with more than 400 organisations in over 100 countries What plants and fungi occur to deliver this mission. The knowledge obtained 1 on Earth and how is this from this research is disseminated in a number diversity distributed? p2 of different ways from annual reports (e.g. stateoftheworldsplants.org) and web-based What drivers and processes portals (e.g. plantsoftheworldonline.org) to 2 underpin global plant and academic papers. fungal diversity? p32 In the academic year 2017-2018, Kew scientists, in collaboration with numerous What plant and fungal diversity is national and international research partners, 3 under threat and what needs to be published 358 papers in international peer conserved to provide resilience reviewed journals and books. Here we bring to global change? p54 together the abstracts of some of these papers. Due to space constraints we have Which plants and fungi contribute to included only those which are led by a Kew 4 important ecosystem services, scientist; a full list of publications, however, can sustainable livelihoods and natural be found at kew.org/publications capital and how do we manage them? p72 * Indicates Kew staff or research associate authors. -

Simarouba Monograph 3/1/04 Simarouba Amara, Glauca
Simarouba Monograph 3/1/04 Simarouba amara, glauca Family: Simaroubaceae Synonyms: Quassia simarouba, Zwingera amara, Picraena officinalis, Simarouba medicinalis Other Common Names: Aceituno, bitterwood, bois blanc, bois amer, bois frene, bois negresse, caixeta, cajú-rana, cedro blanco daguilla, dysentery bark, gavilan, malacacheta, marubá, marupá, negrito, palo blanco, palo amargo, paradise tree, pitomba, robleceillo, simaba. Overview Botanical Description Simarouba is indigenous to the Amazon rainforest and other tropical areas in Mexico, Cuba, Haita, Jamaica and Central America. It grows up to 20 m high and has a trunk 50 to 80 cm in diameter. It produces bright green leaves 20 to 50 cm in length, small white flowers, and small red fruit. Ethnobotanical Uses In traditional herbal medicine systems the bark, wood and leaves of simarouba have been used for their amoebicide, analgesic, anthelmintic, antibacterial, antidysenteric, antimalarial, antimicrobial, astringent, febrifuge, stomachic, sudorific, tonic and vermifuge properties. The traditional use of graviola has been recorded in herbal medicine systems in the following countries: Guiana,1 Belize,2 Brazil,3-6 Cuba,7,8 French Guyana,9 Haiti10 and Peru.11 Summary of Traditional Uses of Simarouba:12 Bark: Anemia, anorexia, bitter,diarrhea, dysentery, dyspepsia, emmenagogue, fever, hemorrhages, internal bleeding, intestinal worms, malaria, skin sores, sores, stomach and bowel disorders, tonic, wounds. Leaf: Astringent, colitis, diarrhea, digestive, dysentery, emmenogogue,intestinal worms, malaria, skin affections. Root: Diarrhea, dysentery, flatulence, intestinal worms, malaria, stomach pain, tonic. Primary Uses Internal A bark tea is primarily used as the first line of defense for amebic dysentery and diarrhea. It’s also used for viruses.13 External The bark has been traditionally used in herbal medicine systems externally for wounds and skin sores.2,7 © Copyrighted 2004. -

Woody-Tissue Respiration for Simarouba Amara and Minquartia Guianensis, Two Tropical Wet Forest Trees with Different Growth Habits
Oecologia (1994) 100:213-220 9Springer-Verlag 1994 Michael G. Ryan 9Robert M. Hubbard Deborah A. Clark 9Robert L. Sanford, Jr. Woody-tissue respiration for Simarouba amara and Minquartia guianensis, two tropical wet forest trees with different growth habits Received: 9 February 1994 / Accepted: 26 July 1994 Abstract We measured CO 2 etflux from stems of two maintenance respiration was 54% of the total CO2 efflux tropical wet forest trees, both found in the canopy, but for Simarouba and 82% for Minquartia. For our sam- with very different growth habits. The species were ple, sapwood volume averaged 23% of stem volume Simarouba amara, a fast-growing species associated when weighted by tree size, or 40% with no size weight- with gaps in old-growth forest and abundant in sec- ing. Using these fractions, and a published estimate of ondary forest, and Minquartia guianensis, a slow-grow- aboveground dry-matter production, we estimate the ing species tolerant of low-light conditions in annual cost of woody tissue respiration for primary old-growth forest. Per unit of bole surface, CO2 efflux forest at La Selva to be 220 or 350 gCm-2year -1, averaged 1.24 gmol m -2 s-1 for Simarouba and depending on the assumed sapwood volume. These 0.83gmolm-2s -1 for Minquartia. C02 elttux was costs are estimated to be less than 13% of the gross highly correlated with annual wood production (r 2= production for the forest. 0.65), but only weakly correlated with stem diameter (r 2 = 0.22). We also partitioned the CO2 efttux into the Key words Woody tissue respiration" Maintenance functional components of construction and mainte- respiration 9Tropical wet forest trees 9Carbon balance nance respiration. -
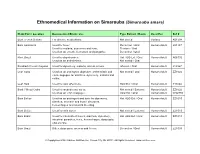
Ethnomedical Information on Simarouba (Simarouba Amara)
Ethnomedical Information on Simarouba (Simarouba amara) Plant Part / Location Documented Ethnic Use Type Extract / Route Used For Ref # Bark French Guiana For diverse medications. Not stated Various K01504 Bark Amazonia Used for fever. Decoction / Oral Human Adult L04137 Used for malaria, dysentery and tonic. Tincture / Oral Used as an emetic, hemostat, and purgative. Decoction / Oral Root Brazil Used to stop diarrhea. Hot H2O Ext / Oral Human Adult A06732 Used as an anthelmintic. Not stated / Oral Rootbark French Guyana Used for dysentery, malaria, and as a tonic. Infusion / Oral Human Adult J12967 Leaf Cuba Used as an astringent, digestive, anthelmintic and Not stated / Oral Human Adult ZZ1022 emmenogogue for diarrhea, dysentery, malaria and colitis. Leaf Haiti Used for skin affections. H2O Ext / Oral Human Adult T13846 Bark / Wood Cuba Used for wounds and sores. Not stated / External Human Adult ZZ1022 Used as an emmenagogue. H2O Ext / Oral Human Adult W02855 Bark Belize Used as an astringent and tonic for dysentery, Hot H2O Ext / Oral Human Adult ZZ1019 diarrhea, stomach and bowel disorders, hemorrhages and internal bleeding. Bark Belize Used for skin sores. Not stated / External Human Adult ZZ1019 Bark Brazil Used for intermittent fevers, diarrhea, dysentery, Hot H2O Ext / Oral Human Adult ZZ1013 intestinal parasites, tonic, hemorrhages, dyspepsia, and anemia. Bark Brazil Bitter, dyspepsia, anemia and fevers. Decoction / Oral Human Adult ZZ1099 © Copyrighted 2004. Raintree Nutrition, Inc. Carson City, NV 89701. All Rights Reserved. www.rain-tree.com Plant Part / Location Documented Ethnic Use Type Extract / Route Used For Ref # Bark + Root Brazil Used for bleeding diarrhea. Decoction / Oral Human Adult ZZ1099 Bark Brazil Used for acute and chronic dysentery, bitter tonic, Infusion / Oral Human Adult ZZ1007 febrifuge, diarrhea, inappetite, and dyspepsia. -

Amazon Plant List
Amazon Plant List The Plant list below is contributed by Dr.Christopher Dick, PhD who has worked in Amazonia for many years. Note that it is a working list and neither exhaustive nor complete. English Common Portuguese Common Plant Family Name Botanical Name Name Name Annonaceae Guatteria Envira-bobô recurvisepala Unonopsis guatterioides Myristicaceae Virola calophylla Wild nutmeg Ucuuba Iryanthera uleii Dead-bark Osteophloeum Ucuuba-amarela platyspermum Lauraceae Mezilaurus itauba Itaúba Persea americana Avocado Abacate Aniba canella Casca preciosa Aniba roseadora Pau rosa Ocotea rubra Louro-gamela Peperomia Piperaceae Ant-garden macrostachya Nymphaeaceae Victoria amazonica Amazon-lily Victoria-regia Menispermaceae Ulmaceae Trema micrantha Trema, Periquitinho Moraceae Clarisia racemosa Guariúba Naucleopsis Miratinga, Pau pica caloneura Brosimim Amapá parinarioides Cecropia Cecropiaceae Purple cecropia Imbaúba roxa purpurascens Cecropia sciadophylla Cecropia Imbaúba-torém Caruru-bravo, Bredo- Phytolaccaceae Phytolacca rivinoides Pokeweed roxo Epiphyllum Cactaceae Cactus phyllanthus Polygonaceae Coccoloba spp. Water-grape? Symeria paniculata Carauaçuzeiro Tetracera Dilleniaceae Water-vine Cipó d'agua willdenowiana Pinzona coriaceae Fire-vine Cipó-de-fôgo Caryocaraceae Caryocar villosum Piquiá Caryocar glabrum Piquiarana Margraviaceae Marcgravia Quiinaceae Clusiaceae Vismia cayennensis Lacre-branco Vismia guianensis Lacre-vermelho Symphonia Ananí used for cerol? globulifera Elaeocarpaceae Sterculiaceae Sterculia frondosa Tacacá Waltheria -

Reconstructing the Deep-Branching Relationships of the Papilionoid Legumes
SAJB-00941; No of Pages 18 South African Journal of Botany xxx (2013) xxx–xxx Contents lists available at SciVerse ScienceDirect South African Journal of Botany journal homepage: www.elsevier.com/locate/sajb Reconstructing the deep-branching relationships of the papilionoid legumes D. Cardoso a,⁎, R.T. Pennington b, L.P. de Queiroz a, J.S. Boatwright c, B.-E. Van Wyk d, M.F. Wojciechowski e, M. Lavin f a Herbário da Universidade Estadual de Feira de Santana (HUEFS), Av. Transnordestina, s/n, Novo Horizonte, 44036-900 Feira de Santana, Bahia, Brazil b Royal Botanic Garden Edinburgh, 20A Inverleith Row, EH5 3LR Edinburgh, UK c Department of Biodiversity and Conservation Biology, University of the Western Cape, Modderdam Road, \ Bellville, South Africa d Department of Botany and Plant Biotechnology, University of Johannesburg, P. O. Box 524, 2006 Auckland Park, Johannesburg, South Africa e School of Life Sciences, Arizona State University, Tempe, AZ 85287-4501, USA f Department of Plant Sciences and Plant Pathology, Montana State University, Bozeman, MT 59717, USA article info abstract Available online xxxx Resolving the phylogenetic relationships of the deep nodes of papilionoid legumes (Papilionoideae) is essential to understanding the evolutionary history and diversification of this economically and ecologically important legume Edited by J Van Staden subfamily. The early-branching papilionoids include mostly Neotropical trees traditionally circumscribed in the tribes Sophoreae and Swartzieae. They are more highly diverse in floral morphology than other groups of Keywords: Papilionoideae. For many years, phylogenetic analyses of the Papilionoideae could not clearly resolve the relation- Leguminosae ships of the early-branching lineages due to limited sampling. -

26 Extreme Trees Pub 2020
Publication WSFNR-20-22C April 2020 Extreme Trees: Tallest, Biggest, Oldest Dr. Kim D. Coder, Professor of Tree Biology & Health Care / University Hill Fellow University of Georgia Warnell School of Forestry & Natural Resources Trees have a long relationship with people. They are both utility and amenity. Trees can evoke awe, mysticism, and reverence. Trees represent great public and private values. Trees most noticed and celebrated by people and communities are the one-tenth of one-percent of trees which approach the limits of their maximum size, reach, extent, and age. These singular, historic, culturally significant, and massive extreme trees become symbols and icons of life on Earth, and our role model in environmental stewardship and sustainability. What Is A Tree? Figure 1 is a conglomeration of definitions and concepts about trees from legal and word definitions in North America. For example, 20 percent of all definitions specifically state a tree is a plant. Concentrated in 63% of all descriptors for trees are four terms: plant, woody, single stem, and tall. If broad stem diameter, branching, and perennial growth habit concepts are added, 87% of all the descriptors are represented. At its most basic level, defining a tree is not species based, but is a structural definition. A tree is represented by a type of plant architecture recognizable by non-technical people. The most basic concepts for defining a tree are — a large, tall, woody, perennial plant with a single, unbranched, erect, self-supporting stem holding an elevated and distinct crown of branches, and which is greater than 10 feet in height and greater than 3 inches in diameter.