How an Improved Sorghum Variety Evolves in a Traditional Seed System in Mali: Effects of Farmers’ Practices on the Maintenance of Phenotype and Genetic Composition
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FINAL REPORT Quantitative Instrument to Measure Commune
FINAL REPORT Quantitative Instrument to Measure Commune Effectiveness Prepared for United States Agency for International Development (USAID) Mali Mission, Democracy and Governance (DG) Team Prepared by Dr. Lynette Wood, Team Leader Leslie Fox, Senior Democracy and Governance Specialist ARD, Inc. 159 Bank Street, Third Floor Burlington, VT 05401 USA Telephone: (802) 658-3890 FAX: (802) 658-4247 in cooperation with Bakary Doumbia, Survey and Data Management Specialist InfoStat, Bamako, Mali under the USAID Broadening Access and Strengthening Input Market Systems (BASIS) indefinite quantity contract November 2000 Table of Contents ACRONYMS AND ABBREVIATIONS.......................................................................... i EXECUTIVE SUMMARY............................................................................................... ii 1 INDICATORS OF AN EFFECTIVE COMMUNE............................................... 1 1.1 THE DEMOCRATIC GOVERNANCE STRATEGIC OBJECTIVE..............................................1 1.2 THE EFFECTIVE COMMUNE: A DEVELOPMENT HYPOTHESIS..........................................2 1.2.1 The Development Problem: The Sound of One Hand Clapping ............................ 3 1.3 THE STRATEGIC GOAL – THE COMMUNE AS AN EFFECTIVE ARENA OF DEMOCRATIC LOCAL GOVERNANCE ............................................................................4 1.3.1 The Logic Underlying the Strategic Goal........................................................... 4 1.3.2 Illustrative Indicators: Measuring Performance at the -

Dossier Technique Et Financier
DOSSIER TECHNIQUE ET FINANCIER PROJET D’APPUI AUX INVESTISSEMENTS DES COLLECTIVITES TERRITORIALES MALI CODE DGD : 3008494 CODE NAVISION : MLI 09 034 11 TABLE DES MATIÈRES ABRÉVIATIONS ............................................................................................................................ 4 RÉSUMÉ ....................................................................................................................................... 6 FICHE ANALYTIQUE DE L’INTERVENTION ............................................................................... 8 1 ANALYSE DE LA SITUATION .............................................................................................. 9 1.1 STRATÉGIE NATIONALE .......................................................................................................... 9 1.2 L’IMPACT DE LA CRISE .......................................................................................................... 11 1.3 DISPOSITIF INSTITUTIONNEL DE LA DÉCENTRALISATION ET LES DISPOSITIFS D’APPUI À LA MISE EN ŒUVRE DE LA RÉFORME ................................................................................................................. 12 1.4 L’ANICT ............................................................................................................................ 15 1.5 QUALITÉ DES INVESTISSEMENTS SOUS MAÎTRISE D’OUVRAGE DES CT .................................... 25 1.6 CADRE SECTORIEL DE COORDINATION, DE SUIVI ET DE DIALOGUE ........................................... 29 1.7 CONTEXTE DE -

Annuaire Statistique 2015 Du Secteur Développement Rural
MINISTERE DE L’AGRICULTURE REPUBLIQUE DU MALI ----------------- Un Peuple - Un But – Une Foi SECRETARIAT GENERAL ----------------- ----------------- CELLULE DE PLANIFICATION ET DE STATISTIQUE / SECTEUR DEVELOPPEMENT RURAL Annuaire Statistique 2015 du Secteur Développement Rural Juin 2016 1 LISTE DES TABLEAUX Tableau 1 : Répartition de la population par région selon le genre en 2015 ............................................................ 10 Tableau 2 : Population agricole par région selon le genre en 2015 ........................................................................ 10 Tableau 3 : Répartition de la Population agricole selon la situation de résidence par région en 2015 .............. 10 Tableau 4 : Répartition de la population agricole par tranche d'âge et par sexe en 2015 ................................. 11 Tableau 5 : Répartition de la population agricole par tranche d'âge et par Région en 2015 ...................................... 11 Tableau 6 : Population agricole par tranche d'âge et selon la situation de résidence en 2015 ............. 12 Tableau 7 : Pluviométrie décadaire enregistrée par station et par mois en 2015 ..................................................... 15 Tableau 8 : Pluviométrie décadaire enregistrée par station et par mois en 2015 (suite) ................................... 16 Tableau 9 : Pluviométrie enregistrée par mois 2015 ........................................................................................ 17 Tableau 10 : Pluviométrie enregistrée par station en 2015 et sa comparaison à -

Bureau Du PAM Au Mali Achat Pour Le Progrès Etudes De Cas Rapport
Bureau du PAM au Mali Achat Pour le Progrès Etudes de Cas Rapport Final Consolidé Bamako, Décembre 2012 P4P Mali Case studies report Page 1 Table des matières Introduction ........................................................................................................................................ 6 Méthodologie ..................................................................................................................................... 6 1. Etudes de cas Producteurs .......................................................................................................... 9 1.1. Un jeune agriculteur leader et membre d’un ménage ............................................................. 9 1.2. Une Agricultrice Membre d’un Ménage dirigé par un Homme ................................................ 13 1.3. Un agriculteur chef de ménage .......................................................................................... 17 1.4. Un agriculteur leader et chef de ménage ............................................................................. 21 1.5. Une agricultrice membre d’un ménage dirigé par un homme ................................................. 24 1.6. De la production d’arachide à la production de niébé : le cas de Mariam Sara, une agricultrice leader et membre d’un ménage dirigé par un homme ....................................................................... 27 1.7. Une agricultrice chef de ménage ........................................................................................ 31 1.8. Un -

Régions De SEGOU Et MOPTI République Du Mali P! !
Régions de SEGOU et MOPTI République du Mali P! ! Tin Aicha Minkiri Essakane TOMBOUCTOUC! Madiakoye o Carte de la ville de Ségou M'Bouna Bintagoungou Bourem-Inaly Adarmalane Toya ! Aglal Razelma Kel Tachaharte Hangabera Douekiré ! Hel Check Hamed Garbakoira Gargando Dangha Kanèye Kel Mahla P! Doukouria Tinguéréguif Gari Goundam Arham Kondi Kirchamba o Bourem Sidi Amar ! Lerneb ! Tienkour Chichane Ouest ! ! DiréP Berabiché Haib ! ! Peulguelgobe Daka Ali Tonka Tindirma Saréyamou Adiora Daka Salakoira Sonima Banikane ! ! Daka Fifo Tondidarou Ouro ! ! Foulanes NiafounkoéP! Tingoura ! Soumpi Bambara-Maoude Kel Hassia Saraferé Gossi ! Koumaïra ! Kanioumé Dianké ! Leré Ikawalatenes Kormou © OpenStreetMap (and) contributors, CC-BY-SA N'Gorkou N'Gouma Inadiatafane Sah ! ! Iforgas Mohamed MAURITANIE Diabata Ambiri-Habe ! Akotaf Oska Gathi-Loumo ! ! Agawelene ! ! ! ! Nourani Oullad Mellouk Guirel Boua Moussoulé ! Mame-Yadass ! Korientzé Samanko ! Fraction Lalladji P! Guidio-Saré Youwarou ! Diona ! N'Daki Tanal Gueneibé Nampala Hombori ! ! Sendegué Zoumané Banguita Kikara o ! ! Diaweli Dogo Kérengo ! P! ! Sabary Boré Nokara ! Deberé Dallah Boulel Boni Kérena Dialloubé Pétaka ! ! Rekerkaye DouentzaP! o Boumboum ! Borko Semmi Konna Togueré-Coumbé ! Dogani-Beré Dagabory ! Dianwely-Maoundé ! ! Boudjiguiré Tongo-Tongo ! Djoundjileré ! Akor ! Dioura Diamabacourou Dionki Boundou-Herou Mabrouck Kebé ! Kargue Dogofryba K12 Sokora Deh Sokolo Damada Berdosso Sampara Kendé ! Diabaly Kendié Mondoro-Habe Kobou Sougui Manaco Deguéré Guiré ! ! Kadial ! Diondori -

Region De Koulikoro
Répartition par commune de la population résidente et des ménages Taux Nombre Nombre Nombre Population Population d'accroissement de de d’hommes en 2009 en 1998 annuel moyen ménages femmes (1998-2009) Cercle de Kati Tiele 2 838 9 220 9 476 18 696 14 871 2,1 Yelekebougou 1 071 3 525 3 732 7 257 10 368 -3,2 Cercle de Kolokani REGION DE KOULIKORO Kolokani 7 891 27 928 29 379 57 307 33 558 5,0 Didieni 4 965 17 073 17 842 34 915 25 421 2,9 En 2009, la région de Koulikoro compte 2 418 305 habitants répartis dans 366 811 ména- ème Guihoyo 2 278 8 041 8 646 16 687 14 917 1,0 ges, ce qui la place au 2 rang national. La population de Koulikoro est composée de Massantola 5 025 17 935 17 630 35 565 29 101 1,8 1 198 841 hommes et de 1 219 464 femmes, soit 98 hommes pour 100 femmes. Les fem- Nonkon 2 548 9 289 9 190 18 479 14 743 2,1 mes représentent 50,4% de la population contre 49,6% pour les hommes. Nossombougou 2 927 10 084 11 028 21 112 17 373 1,8 La population de Koulikoro a été multipliée par près de 1,5 depuis 1998, ce qui représente Ouolodo 1 462 4 935 5 032 9 967 9 328 0,6 un taux de croissance annuel moyen de 4%. Cette croissance est la plus importante jamais Sagabala 2 258 7 623 8 388 16 011 15 258 0,4 constatée depuis 1976. -

M600kv1905mlia1l-Mliadm22303-Sikasso.Pdf (Français)
! ! ! ! ! ! ! ! ! ! ! RÉGION DE SIKASSO - MALI ! ! Map No: MLIADM22303 N'Ga!ssola Yang!asso ! ! ! ! 8°0'W Keninkou 7°0'W Diaforongo 6°0'W 5°0'W Koulala Sourountouna Sanékuy Torodo Baraouéli Diéna ! ! ! Kazangasso ! Niamana Sobala Diéli ! N N ' Mandiakuy ' 0 0 ° ° 3 3 1 Bossofola Yéelekebougou ! 1 ! Kal!aké ! Tiémena Kéme!ni ! Sanando Niala Bla Kalifabougou ! ! Nafadie ! ! Dougouolo ! Karaba Kagoua ! Konobougou Falo Dioundiou Baoulé Diakourouna Nerissso ! ! ! Konkankan ! Diora Koulikoro Gouni S É G O U Somasso Samabogo ! RÉGINOég!ueNla DE SIKASSO P ! Waki 1 ! ! ! Negala Guinina Diaramana Kimparana Begu! ené ^ National Capital Route Pr!incipale ! ! Kambila Dio Gare Gouendo ! M!afouné ! Fana ! Diago ! Safo Talikourou P Chef-lieu Région Rou!te Secondaire Tingolé ! Nianaso! ! Koloni Dombila Kati Kérela Toko!unko ! ! Tienfala ! ! Debéla Baramana Chef-lieu Cercle Tertiary ! ! ! Toura-Kalanga Dialakorodji ! ! Marka-Coungo Kia ! ! ! Chef-lieu Commune FrDoonutibèarbeo Iungteorunationale ! ! ! Tyenfala Sountiani M'Pebougou Pegu!éna Fonfona ! ! Nangola ! ! Kore ! ! ! ! ! ! ! B!ongosso Moribila-Kagoua Kinikai Village Limite Région ! ! ! Toukoro M'Pessoba Zoumanabougou ^ Miéna Dogodouma ! ! Limite Cercle ! M'Pedougou Kenyebaoulé Aéroport ! ! Zan!soni 7 Gouala!bougou ! Wacoro N'Tagonasso ! ! ! M’Pessoba Ou!la ! Faya ! Tasso Fleuve Bamako Baramba ! Koumbia ! ! Sirababougou ! Zone Marécageuse ! Karagouana Mallé! Daboni ! Dandougou N'Tongo!losso ! Kouniana N'Pa!nafa Nien!esso Forêts Classées ! Ouenzzindougou ! K O U L I K O R O ! Bobola-Zangasso ! Lac Kolomosso ! Dorokouma ! Torosogoba Sinde ! 7 ! Bamana N'Tossoni Sorob!asso Djigo!uala Sirake!lé Mountougoula ! ! Ko!un Bele!koro ! Kiffosso 1 Beleko Soba Togobabougou Zangorola Cette carte a été réalisée selon le découpage adminisMtraotnift sd uM Maanldi iàn pgauretisr des ! ! Kon!ina ! Hamidou Goita Dioila ! Gouama Sogotila Famessasso Beledo!ugou ! données de la Direction Nationale des Collectivités Territoriales (DNCT). -

First Mali Report on the Implementation of the African Charter on the Rights and Welfare of the Child
Republic of Mali One People – One Goal – One Faith FIRST MALI REPORT ON THE IMPLEMENTATION OF THE AFRICAN CHARTER ON THE RIGHTS AND WELFARE OF THE CHILD FOR THE 1999 – 2006 PERIOD Bamako, September 2007 ACRONYMS AND ABBREVIATIONS AN-RM National Assembly of the Republic of Mali. PTA Parents-Teachers Association ARV Anti- Retroviral ASACO Association de Santé Communautaire (Community Health Association) CAFO Coordination of Associations and Women NGOs of Mali CRC Convention on the Rights of the Child CED Centre d’Education pour le Développement (Education Centre for Development) CESC Economic, Social and Cultural Council CNAPN Comité National d’Action pour l’Eradication des Pratiques Néfastes à la santé de la Femme et de l’Enfant (National Action Committee for the Eradication of Harmful Practices on the Health of the Women and the Child) CNDIFE Centre National de Documentation et d’Information sur la Femme et l’Enfant. (National Documentation and Information Centre on the Woman and the Child) COMADE Coalition Malienne des Droits de l’Enfant (Malian Coalition for the Right of the Child) CPE Code for the Protection of the Child CSCOM Centre de Santé Communautaire (Community Health Centre) CSLP Poverty Alleviation Strategic Framework DNPF National Department of Women Empowerment. DNPEF National Department for the Promotion of the Child and the Family DNSI National Department of Statistics and Data Processing EDS III Population and Health Survey of Mali 2001 HCCT High Council of Territorial Councils IMAARV Malian Initiative for Access to Anti-Retrovirals -

Rgion De Koulikoro Rpublique Du Mali
PRESIDENCE DE LA REPUBLIQUE REPUBLIQUE DU MALI ---------------------- Un Peuple – Un But – Une Foi Commissariat à la Sécurité Alimentaire (CSA) ----------------------- Projet de Mobilisation des Initiatives en matière de Sécurité Alimentaire au Mali (PROMISAM) REGION DE Koulikoro Cercle de Dioila Commune rurale de Wacoro PLAN DE SECURITE ALIMENTAIRE COMMUNE RURALE DE WACORO 2008- 2012 Elaboré avec l’appui technique et financier de l’USAID-Mali à travers le projet d’appui au CSA, le PROMISAM Avril 2008 I PRESENTATION DE LA COMMUNE La Commune de Wacoro fait partie des six (6) communes de l’arrondissement central de Dioila. 1-1- Historique : Le village de Wacoro serait fondé au 16e siècle par Ndjda COULIBALY ressortissant de Bana dans l’actuel cercle de Bouguini le premier site du village serait N’DJBALA puis Siriki, il serait donc à son 3e site. Il a successivement fait face aux agressions des mois Bamanan de Ségou, de SAMORY TOURE du Wassoulou et d’El Hadji Oumar TALL de l’empire TOUCOULEUR. Le village a connu de braves guerriers comme Bala Coulibaly dit Wacoro Baba et Nama Coulibaly –Hostile à l’administration coloniale, Wacoro a été rattaché au canton de Nangola. En 1960, Wacoro a été rattaché à l’arrondissement central de Dioila dont il était le 2e village par la taille de sa population 1-2-Situation Géographique : Située à 25 km au Nord –Est de Dioila, la commune de Wacoro a été créée par la loi N°96 – 059 / AN.RM du 4/4/1996 –Elle est limitée : • à l’Ouest par les communes de Kérela et Kaladougou • au Nord – est par la commune de Nangola • au Sud par le fleuve Baoulé qui le sépare de Kilidougou et de Kaladougou. -

GE84/210 BR IFIC Nº 2747 Section Spéciale Special Section Sección
Section spéciale Index BR IFIC Nº 2747 Special Section GE84/210 Sección especial Indice International Frequency Information Circular (Terrestrial Services) ITU - Radiocommunication Bureau Circular Internacional de Información sobre Frecuencias (Servicios Terrenales) UIT - Oficina de Radiocomunicaciones Circulaire Internationale d'Information sur les Fréquences (Services de Terre) UIT - Bureau des Radiocommunications Date/Fecha : 25.06.2013 Expiry date for comments / Fecha limite para comentarios / Date limite pour les commentaires : 03.10.2013 Description of Columns / Descripción de columnas / Description des colonnes Intent Purpose of the notification Propósito de la notificación Objet de la notification 1a Assigned frequency Frecuencia asignada Fréquence assignée 4a Name of the location of Tx station Nombre del emplazamiento de estación Tx Nom de l'emplacement de la station Tx B Administration Administración Administration 4b Geographical area Zona geográfica Zone géographique 4c Geographical coordinates Coordenadas geográficas Coordonnées géographiques 6a Class of station Clase de estación Classe de station 1b Vision / sound frequency Frecuencia de portadora imagen/sonido Fréquence image / son 1ea Frequency stability Estabilidad de frecuencia Stabilité de fréquence 1e carrier frequency offset Desplazamiento de la portadora Décalage de la porteuse 7c System and colour system Sistema de transmisión / color Système et système de couleur 9d Polarization Polarización Polarisation 13c Remarks Observaciones Remarques 9 Directivity Directividad -
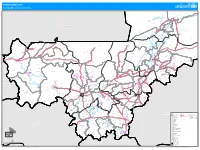
PROGRAMME ATPC Cartographie Des Interventions
PROGRAMME ATPC Cartographie des interventions Agouni !( Banikane !( TOMBOUCTOU Rharous ! Ber . !( Essakane Tin AÎcha !( Tombouctou !( Minkiri Madiakoye H! !( !( Tou!(cabangou !( Bintagoungou M'bouna Bourem-inaly !( !( Adarmalane Toya !( !( Aglal Raz-el-ma !( !( Hangabera !( Douekire GOUNDAM !( Garbakoira !( Gargando Dangha !( !( G!(ou!(ndam Sonima Doukouria Kaneye Tinguereguif Gari !( .! !( !( !( !( Kirchamba TOMBOUCTOU !( MAURITANIE Dire .! !( HaÏbongo DIRE !( Tonka Tindirma !( !( Sareyamou !( Daka Fifo Salakoira !( GOURMA-RHAROUS Kel Malha Banikane !( !( !( NIAFOUNKE Niafunke .! Soumpi Bambara Maoude !( !( Sarafere !( KoumaÏra !( Dianke I Lere !( Gogui !( !( Kormou-maraka !( N'gorkou !( N'gouma Inadiatafane Sah !( !( !( Ambiri !( Gathi-loumo !( Kirane !( Korientze Bafarara Youwarou !( Teichibe !( # YOUWAROU !( Kremis Guidi-sare !( Balle Koronga .! !( Diarra !( !( Diona !( !( Nioro Tougoune Rang Gueneibe Nampala !( Yerere Troungoumbe !( !( Ourosendegue !( !( !( !( Nioro Allahina !( Kikara .! Baniere !( Diaye Coura !( # !( Nara Dogo Diabigue !( Gavinane Guedebin!(e Korera Kore .! Bore Yelimane !( Kadiaba KadielGuetema!( !( !( !( Go!(ry Youri !( !( Fassoudebe Debere DOUENTZA !( .! !( !( !(Dallah Diongaga YELIMANE Boulal Boni !( !(Tambacara !( !( Takaba Bema # # NIORO !( # Kerena Dogofiry !( Dialloube !( !( Fanga # Dilly !( !( Kersignane !( Goumbou # KoubewelDouentza !( !( Aourou !( ## !( .! !( # K#onna Borko # # #!( !( Simbi Toguere-coumbe !( NARA !( Dogani Bere Koussane # !( !( # Dianwely-maounde # NIONO # Tongo To !( Groumera Dioura -

World Bank Document
Document of The World Bank FOR OFFICIAL USE ONLY Public Disclosure Authorized Report No: 32130 IMPLEMENTATION COMPLETION REPORT (IDA-26170) ON A CREDIT Public Disclosure Authorized IN THE AMOUNT OF SDR 46.1 MILLION (US$ 65 MILLION EQUIVALENT) TO THE REPUBLIC OF MALI FOR A TRANSPORT SECTOR PROJECT Public Disclosure Authorized June 20, 2005 Transport Sector Country Department 15 Africa Region This document has a restricted distribution and may be used by recipients only in the performance of their Public Disclosure Authorized official duties. Its contents may not otherwise be disclosed without World Bank authorization. CURRENCY EQUIVALENTS (Exchange Rate Effective May 5, 1994 - December 31, 2004) Currency Unit = CFA Franc (CFAF) 1CFAF = US$ 0.0016 US$ 1.00 = CFAF 600 US$ 1.00 = CFAF 602.4 (average during project life) FISCAL YEAR January 1 to December 31 ABBREVIATIONS AND ACRONYMS ABEDA - Arab Bank for Economic Development in Africa ADM - Aéroports du Mali (Mali Airports) ASECNA - Agency for the Safety of Aerial Navigation in Africa and Madagascar BOAD - Banque Ouest Africaine de Développement (West African Development Bank) CEAO - Communauté Economique de l’Afrique de l'Ouest (West African Economic Community) CFD - Caisse Française de Développement (French Development Fund) CIDA - Canadian International Development Agency CIF - Cost, Insurance and Freight (border point price) CMDT - Compagnie Malienne de Développement des Textiles (Malian Textile Development Company) CNREX - Centre National de Recherche et d'Expérimentation pour le Bâtiment