The Understudied and Underappreciated Role of Predation in the Mortality of fish Released from fishing Gears
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Traditional Ecological Knowledge in Andavadoaka, Southwest Madagascar
BLUE VENTURES CONSERVATION REPORT Josephine M. Langley Vezo Knowledge: Traditional Ecological Knowledge in Andavadoaka, southwest Madagascar 52 Avenue Road, London N6 5DR [email protected] Tel: +44 (0) 208 341 9819 Fax: +44 (0) 208 341 4821 BLUE VENTURES CONSERVATION REPORT Andavadoaka, 2006 © Blue Ventures 2006 Copyright in this publication and in all text, data and images contained herein, except as otherwise indicated, rests with Blue Ventures. Recommended citation: Langley, J. (2006). Vezo Knowledge: Traditional Ecological Knowledge in Andavadoaka, southwest Madagascar. BLUE VENTURES CONSERVATION REPORT Summary Many fisheries and marine conservation management (ii) Knowledge of marine resources is passed orally from projects throughout the world have been dogged by generation to generation failure as a result of a lack of acceptance of (iii) Traditional laws, taboos and ceremonies are commonly management interventions by local communities. used in natural resource management Evaluation of these failed studies has produced (iv) Lifestyle, food security and housing are all dependent extensive guidelines, manuals and new fields of study on natural resources and the use of coastal and marine (Bunce and Pomeroy, 2004). Community engagement, resources form an essential part of this participatory research, and promoting the use of local (v) The arrival of the Catholic Mission has reduced the knowledge have repeatedly emerged as steps necessary proportion of villagers adhering to traditional ancestor to address the problem of managing the development of worship people and their economies while simultaneously (vi) There has been a change in recent years from a barter protecting the environment (Berkes et al, 2001; Bunce and subsistence economy to a market-driven cash- based economy and Pomeroy, 2004; Wibera et al, 2004; Scholz et al, (vii) Increased income for some members of the 2004). -

Marine Biodiversity of the South East NRM Region
Marine Environment and Ecology Benthic Ecology Subprogram Marine Biodiversity of the South East NRM Region SARDI Publication No. F2009/000681-1 SARDI Research Report series No. 416 Keith Rowling, Shirley Sorokin, Leonardo Mantilla and David Currie SARDI Aquatic Sciences PO BOX 120 Henley Beach SA 5022 December 2009 Prepared for the Department for Environment and Heritage 1 Information Systems and Database Support Program Marine Biodiversity of the South East NRM Region Keith Rowling, Shirley Sorokin, Leonardo Mantilla and David Currie December 2009 SARDI Publication No. F2009/000681-1 SARDI Research Report Series No. 416 Prepared for the Department for Environment and Heritage 2 This Publication may be cited as: Rowling, K.P., Sorokin, S.J., Mantilla, L. & Currie, D.R.. (2009) Marine Biodiversity of the South East NRM Region. South Australian Research and Development Institute (Aquatic Sciences), Adelaide. SARDI Publication No. F2009/000681-1. South Australian Research and Development Institute SARDI Aquatic Sciences 2 Hamra Avenue West Beach SA 5024 Telephone: (08) 8207 5400 Facsimile: (08) 8207 5406 http://www.sardi.sa.gov.au DISCLAIMER The authors warrant that they have taken all reasonable care in producing this report. The report has been through the SARDI internal review process, and has been formally approved for release by the Chief of Division. Although all reasonable efforts have been made to ensure quality, SARDI does not warrant that the information in this report is free from errors or omissions. SARDI does not accept any liability for the contents of this report or for any consequences arising from its use or any reliance placed upon it. -

Sharkcam Fishes
SharkCam Fishes A Guide to Nekton at Frying Pan Tower By Erin J. Burge, Christopher E. O’Brien, and jon-newbie 1 Table of Contents Identification Images Species Profiles Additional Info Index Trevor Mendelow, designer of SharkCam, on August 31, 2014, the day of the original SharkCam installation. SharkCam Fishes. A Guide to Nekton at Frying Pan Tower. 5th edition by Erin J. Burge, Christopher E. O’Brien, and jon-newbie is licensed under the Creative Commons Attribution-Noncommercial 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc/4.0/. For questions related to this guide or its usage contact Erin Burge. The suggested citation for this guide is: Burge EJ, CE O’Brien and jon-newbie. 2020. SharkCam Fishes. A Guide to Nekton at Frying Pan Tower. 5th edition. Los Angeles: Explore.org Ocean Frontiers. 201 pp. Available online http://explore.org/live-cams/player/shark-cam. Guide version 5.0. 24 February 2020. 2 Table of Contents Identification Images Species Profiles Additional Info Index TABLE OF CONTENTS SILVERY FISHES (23) ........................... 47 African Pompano ......................................... 48 FOREWORD AND INTRODUCTION .............. 6 Crevalle Jack ................................................. 49 IDENTIFICATION IMAGES ...................... 10 Permit .......................................................... 50 Sharks and Rays ........................................ 10 Almaco Jack ................................................. 51 Illustrations of SharkCam -

Biological Assessment of Kahana Stream, Island of O'ahu
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by ScholarSpace at University of Hawai'i at Manoa Biological Assessment of Kahana Stream, Island of O‘ahu, Hawai‘i: An Application of PABITRA Survey Methods1 J. M. Fitzsimons,2 J. E. Parham,3 L. K. Benson,4 M. G. McRae,2 and R. T. Nishimoto5 Abstract: Aquatic biologists surveyed Kahana Stream on O‘ahu, Hawai‘i, during December 2001 and January, March, and May 2002 to provide a background of information before restoring water diverted from the headwaters of the stream since the mid-1920s. Kahana Stream has all but one species of macrofauna com- mon in unaltered Hawaiian streams, but abundance and distribution of am- phidromous species differ conspicuously. A single specimen of ‘o‘opu ‘alamo‘o (Lentipes concolor) was found near the headwaters; until recently, this species was regarded as extinct on O‘ahu. Only two individuals of the freshwater limpet (hı¯hı¯wai, Neritina granosa) were found, and the brackish-water limpet (hapawai, Neritina vespertina) was not observed. Construction of the Waia¯hole Ditch Tun- nel about 80 yr ago reduced the amount of water entering Kahana headwaters, and unimpeded growth of hau (Hibiscus tiliaceus) from the shore into the stream has slowed water movement in the middle and lower sections of the stream and estuary. Reduced flow has resulted in an extension farther inland of certain es- tuarine and lower-reach species (the prawn Macrobrachium grandimanus and fishes Eleotris sandwicensis and Stenogobius hawaiiensis). Alien fishes and larger in- vertebrates occur throughout Kahana Stream. -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

Fish Bulletin No. 37. the California Barracuda (Sphyraena Argentea)
UC San Diego Fish Bulletin Title Fish Bulletin No. 37. The California Barracuda (Sphyraena argentea). I. Life History of the California Barracuda. II. A Bibliography of Barracudas (Sphyraenidae) Permalink https://escholarship.org/uc/item/3dp2c999 Author Walford, Lionel A Publication Date 1931-09-01 eScholarship.org Powered by the California Digital Library University of California DIVISION OF FISH AND GAME OF CALIFORNIA BUREAU OF COMMERCIAL FISHERIES FISH BULLETIN No. 37 The California Barracuda (Sphyraena argentea) By LIONEL A. WALFORD 1 2 3 4 1. LIFE HISTORY OF THE CALIFORNIA BARRACUDA 5 1.1. ACKNOWLEDGMENTS The study of the life history of the barracuda, of which this paper is the report, was initiated by the International Fisheries Commission (United States and Mexico). Most of the preliminary survey of the fishery and the literature was carried on under the direction of this body. The material collected during this period was kindly turned over to the California Division of Fish and Game in 1927 when the United States-Mexican convention was abrogated. Several people have contributed in various ways to the progress of this investigation. Mr. W. L. Scofield, Director of the California State Fisheries Laboratory, was frequently consulted for advice and suggestions. Commercial fish- ermen and dealers generously permitted measurements of fish on the boats and in the markets. Mr. Scotti Carmen of San Pedro and Mr. A. Wagner of Long Beach made possible the collection of young fish for the growth studies. Mrs. Hilda E. Walford aided in tabulating frequency records from the original data. Dr. F. W. Weymouth of Stan- ford University and Dr. -

Final Report
FINAL REPORT REHABILITATION AND CONSERVATION THE SEAGRASS MEADOWS AT CAM HAI DONG, CAM RANH BAY, KHANH HOA PROVINCE, CENTRAL VIETNAM. Pham Huu Tri Institute of Oceanography Nhatrang, Vietnam NHATRANG, JUNE 2008 Table of Contents 1. INTRODUCTION: 3 2. OBJECTIVES: 4 3. MATERIALS AND METHODS: 4 3.1 Research methods: 4 3.2 Project sites: 4 3. 3 Duration of the project: 6 3.4 Planting techniques: 6 3.4.1 Seagrasses: 6 3.4.2 Sea-horse and sea-cucumber: 7 3.4.3 Methods of study the leaf growth rate and leaf production: 7 3.4.4 Management and taking care of the rehabilitated area at Cam Hai Dong, Cam Ranh ba: 7 4. RESULTS: 7 4.1 The natural environmental elements in the rehabilitated area – Cam Hai Dong- Cam Ranh: 7 4.2 Species diversity of seagrasses in the rehabilitated area: 8 4.3 Transplantation the Enhalus acoroides for planting: 9 4.3.1 For the shoots method: 9 4.3.2 For the seedlings methods: 11 4. 4 Results of restoration the sea-horse at the rehabilitated area: 13 4.5 Results of restoration the sea-cucumber at the rehabilitated area: 14 5. CONCLUSION: 15 6. RECOMMENDATION: 15 LITERATURE CITED: 16 APPENDIX: 17 2 1. INTRODUCTION: Khanh Hoa is the coastal province in southern central Vietnam, situated between 11°45’02’’ to 13°02’ 10’’ North latitude and from 108045’10’’to 109045’06’’ East longitude, the length of the coast lines about 300km. The shallow coastal area of Khanh Hoa province is often covered by seagrass beds especially in Cam Ranh bay area where the seagrass beds have concentrated distribution with the superficies up to more than 1000 ha, there are 6 common species of seagrasses have been found there, among them Enhalus acoroides which has long leaves (1-2 m), dense density (40- 100 shoots/m2) and high cover (70-100%) is dominant. -

Proceedings of the American Lobster Transferable Trap Workshop
Special Report No. 75 of the Atlantic States Marine Fisheries Commission Proceedings of the American Lobster Transferable Trap Workshop October 2002 Proceedings of the American Lobster Transferable Trap Workshop October 2002 Edited by Heather M. Stirratt Atlantic States Marine Fisheries Commission Convened by: Atlantic States Marine Fisheries Commission August 26, 2002 Washington, DC Proceedings of the American Lobster Transferable Trap Workshop ii Preface This document was prepared in cooperation with the Atlantic States Marine Fisheries Commission’s American Lobster Management Board, Technical Committee, Plan Development Team, Stock Assessment Subcommittee and the Advisory Panel. A report of the Atlantic States Marine Fisheries Commission pursuant to U.S. Department of Commerce, National Oceanic and Atmospheric Administration Award No. NA17FG2205. Proceedings of the American Lobster Transferable Trap Workshop iii Acknowledgments This report is the result of a Workshop on transferable trap programs for American lobster which was held on August 26, 2002, in Washington, DC. The workshop was convened and organized by a Workshop Steering Committee composed of: David Spencer (Advisory Panel, Chair), George Doll (New York Commercial Lobsterman, Advisory Panel Member), John Sorlien (Rhode Island Commercial Lobsterman, Advisory Panel Member), Todd Jesse (Massachusetts Commercial Lobsterman, Advisory Panel Member), Ernie Beckwith (Connecticut Department of Environmental Protection), Mark Gibson (Rhode Island Department of Environmental Management), -

Intrinsic Vulnerability in the Global Fish Catch
The following appendix accompanies the article Intrinsic vulnerability in the global fish catch William W. L. Cheung1,*, Reg Watson1, Telmo Morato1,2, Tony J. Pitcher1, Daniel Pauly1 1Fisheries Centre, The University of British Columbia, Aquatic Ecosystems Research Laboratory (AERL), 2202 Main Mall, Vancouver, British Columbia V6T 1Z4, Canada 2Departamento de Oceanografia e Pescas, Universidade dos Açores, 9901-862 Horta, Portugal *Email: [email protected] Marine Ecology Progress Series 333:1–12 (2007) Appendix 1. Intrinsic vulnerability index of fish taxa represented in the global catch, based on the Sea Around Us database (www.seaaroundus.org) Taxonomic Intrinsic level Taxon Common name vulnerability Family Pristidae Sawfishes 88 Squatinidae Angel sharks 80 Anarhichadidae Wolffishes 78 Carcharhinidae Requiem sharks 77 Sphyrnidae Hammerhead, bonnethead, scoophead shark 77 Macrouridae Grenadiers or rattails 75 Rajidae Skates 72 Alepocephalidae Slickheads 71 Lophiidae Goosefishes 70 Torpedinidae Electric rays 68 Belonidae Needlefishes 67 Emmelichthyidae Rovers 66 Nototheniidae Cod icefishes 65 Ophidiidae Cusk-eels 65 Trachichthyidae Slimeheads 64 Channichthyidae Crocodile icefishes 63 Myliobatidae Eagle and manta rays 63 Squalidae Dogfish sharks 62 Congridae Conger and garden eels 60 Serranidae Sea basses: groupers and fairy basslets 60 Exocoetidae Flyingfishes 59 Malacanthidae Tilefishes 58 Scorpaenidae Scorpionfishes or rockfishes 58 Polynemidae Threadfins 56 Triakidae Houndsharks 56 Istiophoridae Billfishes 55 Petromyzontidae -

Bite Force in Two Top Predators, the Great Barracuda, <Em>
University of South Florida Scholar Commons Graduate Theses and Dissertations Graduate School 4-3-2009 Bite Force in Two Top Predators, the Great Barracuda, Sphyraena barracuda and Bull Shark Carcharhinus leucas, During Ontogeny María Laura Habegger University of South Florida Follow this and additional works at: https://scholarcommons.usf.edu/etd Part of the American Studies Commons Scholar Commons Citation Habegger, María Laura, "Bite Force in Two Top Predators, the Great Barracuda, Sphyraena barracuda and Bull Shark Carcharhinus leucas, During Ontogeny" (2009). Graduate Theses and Dissertations. https://scholarcommons.usf.edu/etd/1996 This Thesis is brought to you for free and open access by the Graduate School at Scholar Commons. It has been accepted for inclusion in Graduate Theses and Dissertations by an authorized administrator of Scholar Commons. For more information, please contact [email protected]. Bite Force in Two Top Predators, the Great Barracuda, Sphyraena barracuda and Bull Shark Carcharhinus leucas, During Ontogeny. by María Laura Habegger A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science Department of Biology College of Arts and Science University of South Florida Major Professor: Philip J. Motta, Ph.D. Steven Deban, Ph.D. Daniel Huber, Ph.D. Date of Approval: April 3, 2009 Keywords: biomechanics, performance, functional morphology, scaling, ecomorphology © Copyright 2009, María Laura Habegger Dedication To my parents, for their unconditional support and love, for being by my side even from thousands miles away… Acknowledgments I’m sure I will be able to fill this entire space expressing my gratitude to all the people that helped me through these years, but there is one person that made this dream come true and I will never forget his generosity. -
Field Identification Guide to the Living Marine Resources in Kenya
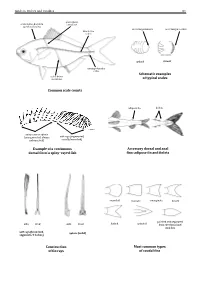
Guide to Orders and Families 81 lateral line scales above scales before dorsal fin outer margin smooth outer margin toothed (predorsal scales) lateral–line 114 scales cycloid ctenoidِّ scales circumpeduncular Schematic examples lateral line of typical scales scales below Common scale counts adipose fin finlets soft rays (segmented, spinyunbranched) rays or spines usually branched) (unsegmented, always Example of a continuous Accessory dorsal and anal dorsal fin of a spiny–rayed fish fins: adipose fin and finlets rounded truncate emarginate lunate side front side front from the dorsal and pointed and separated forked pointed soft rays (branched, spines (solid) segments, 2 halves) anal fins Construction Most common types of fin rays of caudal fins 82 Bony Fishes GUIDE TO ORDERS AND FAMILIES Order ELOPIFORMES – Tarpons and allies Fin spines absent; a single dorsal fin located above middle of body; pelvic fins in abdominal position; lateral line present; 23–25 branchiostegal rays; upper jaw extending past eye; tip of snout not overhanging mouth; colour silvery. ELOPIDAE Page 121 very small scales Ladyfishes To 90 cm. Coastal marine waters and estuaries; pelagic. A single species included in the Guide to Species.underside of head large mouth gular plate MEGALOPIDAE Page 121 last ray long Tarpons large scales To 55 cm. Coastal marine waters and estuaries; pelagic. A single species included in the Guide to Species.underside of head gular plate Order ALBULIFORMES – Bonefishes Fin spines absent; a single dorsal fin located above middle of body; pelvic fins in abdominal position; lateral line present; 6–16 branchiostegal rays; upper jaw not extending as far as front of eye; tip of snout overhanging mouth; colour silvery. -

5-Review-Fish-Habita
United Nations UNEP/GEF South China Sea Global Environment Environment Programme Project Facility UNEP/GEF/SCS/RWG-F.8/5 Date: 12th October 2006 Original: English Eighth Meeting of the Regional Working Group for the Fisheries Component of the UNEP/GEF Project: “Reversing Environmental Degradation Trends in the South China Sea and Gulf of Thailand” Bangka Belitung Province, Indonesia 1st - 4th November 2006 INFORMATION COLLATED BY THE FISHERIES AND HABITAT COMPONENTS OF THE SOUTH CHINA SEA PROJECT ON SITES IMPORTANT TO THE LIFE- CYCLES OF SIGNIFICANT FISH SPECIES UNEP/GEF/SCS/RWG-F.8/5 Page 1 IDENTIFICATION OF FISHERIES REFUGIA IN THE GULF OF THAILAND It was discussed at the Sixth Meeting of the Regional Scientific and Technical Committee (RSTC) in December 2006 that the Regional Working Group on Fisheries should take the following two-track approach to the identification of fisheries refugia: 1. Review known spawning areas for pelagic and invertebrate species, with the aim of evaluating these sites as candidate spawning refugia. 2. Evaluate each of the project’s habitat demonstration sites as potential juvenile/pre-recruit refugia for significant demersal species. Rationale for the Two-Track Approach to the Identification of Fisheries Refugia The two main life history events for fished species are reproduction and recruitment. It was noted by the RSTC that both of these events involve movement between areas, and some species, often pelagic fishes, migrate to particular spawning areas. It was also noted that many species also utilise specific coastal habitats such as coral reefs, seagrass, and mangroves as nursery areas. In terms of the effects of fishing, most populations of fished species are particularly vulnerable to the impacts of high levels of fishing effort in areas and at times where there are high abundances of (a) stock in spawning condition, (b) juveniles and pre-recruits, or (c) pre-recruits migrating to fishing grounds.