Evolution of Oviductal Gestation in Amphibians MARVALEE H
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Kansas 4-H Poultry Leader Notebook Level I Introduction for Leaders
Kansas 4-H Poultry Leader Notebook Level I Introduction for Leaders ........................................................i Parts of a Chicken......................................................................................................3 Name That Bird.......................................................................................................11 Beginning to Set Goals in Your Poultry Project......................................................19 Common Poultry Terms for Different Species........................................................23 Poultry Breeds.........................................................................................................27 Breeds of Poultry for Project and Show..................................................................33 What Bird Will I Raise?..........................................................................................41 Nutritional Needs and Problems in Poultry.............................................................45 Is Your Bird Sick?...................................................................................................51 Catching and Handling Poultry...............................................................................55 Washing That Bird...................................................................................................59 Why Do We Raise Poultry?.....................................................................................63 K-State Research & Extension ■ Manhattan Leader Notes Parts of a Chicken Poultry, -

Great Food, Great Stories from Korea
GREAT FOOD, GREAT STORIE FOOD, GREAT GREAT A Tableau of a Diamond Wedding Anniversary GOVERNMENT PUBLICATIONS This is a picture of an older couple from the 18th century repeating their wedding ceremony in celebration of their 60th anniversary. REGISTRATION NUMBER This painting vividly depicts a tableau in which their children offer up 11-1541000-001295-01 a cup of drink, wishing them health and longevity. The authorship of the painting is unknown, and the painting is currently housed in the National Museum of Korea. Designed to help foreigners understand Korean cuisine more easily and with greater accuracy, our <Korean Menu Guide> contains information on 154 Korean dishes in 10 languages. S <Korean Restaurant Guide 2011-Tokyo> introduces 34 excellent F Korean restaurants in the Greater Tokyo Area. ROM KOREA GREAT FOOD, GREAT STORIES FROM KOREA The Korean Food Foundation is a specialized GREAT FOOD, GREAT STORIES private organization that searches for new This book tells the many stories of Korean food, the rich flavors that have evolved generation dishes and conducts research on Korean cuisine after generation, meal after meal, for over several millennia on the Korean peninsula. in order to introduce Korean food and culinary A single dish usually leads to the creation of another through the expansion of time and space, FROM KOREA culture to the world, and support related making it impossible to count the exact number of dishes in the Korean cuisine. So, for this content development and marketing. <Korean Restaurant Guide 2011-Western Europe> (5 volumes in total) book, we have only included a selection of a hundred or so of the most representative. -

Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca
Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca John F. Lamoreux, Meghan W. McKnight, and Rodolfo Cabrera Hernandez Occasional Paper of the IUCN Species Survival Commission No. 53 Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca John F. Lamoreux, Meghan W. McKnight, and Rodolfo Cabrera Hernandez Occasional Paper of the IUCN Species Survival Commission No. 53 The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN or other participating organizations. Published by: IUCN, Gland, Switzerland Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Citation: Lamoreux, J. F., McKnight, M. W., and R. Cabrera Hernandez (2015). Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca. Gland, Switzerland: IUCN. xxiv + 320pp. ISBN: 978-2-8317-1717-3 DOI: 10.2305/IUCN.CH.2015.SSC-OP.53.en Cover photographs: Totontepec landscape; new Plectrohyla species, Ixalotriton niger, Concepción Pápalo, Thorius minutissimus, Craugastor pozo (panels, left to right) Back cover photograph: Collecting in Chamula, Chiapas Photo credits: The cover photographs were taken by the authors under grant agreements with the two main project funders: NGS and CEPF. -

Management of Neonates Born at ≤34 6/7 Weeks' Gestation with Suspected Or Proven Early-Onset Bacterial Sepsis Karen M
CLINICAL REPORT Guidance for the Clinician in Rendering Pediatric Care ≤ ’ Management of Neonates Born at 34 Karen M. Puopolo, MD, PhD, FAAP, a, b William E. Benitz, MD, FAAP, c Theoklis E. Zaoutis, MD, MSCE, FAAP, a, d 6/7COMMITTEE ONWeeks FETUS AND NEWBORN, GestationCOMMITTEE ON INFECTIOUS DISEASES With Suspected or Proven Early-Onset Bacterial Sepsis Early-onset sepsis (EOS) remains a serious and often fatal illness among abstract infants born preterm, particularly among newborn infants of the lowest gestational age. Currently, most preterm infants with very low birth weight are treated empirically with antibiotics for risk of EOS, often for prolonged aDepartment of Pediatrics, Perelman School of Medicine, University periods, in the absence of a culture-confirmed infection. Retrospective of Pennsylvania, Philadelphia, Pennsylvania; bChildren’s Hospital of studies have revealed that antibiotic exposures after birth are associated Philadelphia, and dRoberts Center for Pediatric Research, Philadelphia, Pennsylvania; and cDivision of Neonatal and Developmental Medicine, with multiple subsequent poor outcomes among preterm infants, making the Department of Pediatrics, School of Medicine, Stanford University, Palo risk/benefit balance of these antibiotic treatments uncertain. Gestational Alto, California age is the strongest single predictor of EOS, and the majority of preterm This document is copyrighted and is property of the American Academy of Pediatrics and its Board of Directors. All authors have births occur in the setting of other factors associated with risk of EOS, filed conflict of interest statements with the American Academy of Pediatrics. Any conflicts have been resolved through a process making it difficult to apply risk stratification strategies to preterm infants. -

Nectophrynoides Tornieri (Anura: Bufonidae) in the Amani Nature Reserve, Tanzania“
DIPLOMARBEIT Titel der Diplomarbeit „The „push-up“ as a calling posture in Nectophrynoides tornieri (Anura: Bufonidae) in the Amani Nature Reserve, Tanzania“ Verfasserin Iris Starnberger angestrebter akademischer Grad Magistra der Naturwissenschaften (Mag.rer.nat.) Wien, 2011 Studienkennzahl lt. A 439 Studienblatt: Studienrichtung lt. Diplomstudium Zoologie (Stzw) UniStG Studienblatt: Betreuerin / Betreuer: Ao. Univ.-Prof. Mag. Dr. Walter Hödl Für meine Eltern und meine Schwester Sonja, die mich immer liebevoll unterstützt und ermutigt haben. 2 Table of Contents INTRODUCTION .............................................................................................................................................4 MATERIALS AND METHODS.......................................................................................................................8 STUDY SITE ......................................................................................................................................................................... 8 EXPERIMENTAL DESIGN ................................................................................................................................................... 8 HABITAT FEATURES....................................................................................................................................................... 10 DATA ANALYSIS ............................................................................................................................................................. -
![Oogenesis [PDF]](https://docslib.b-cdn.net/cover/2902/oogenesis-pdf-452902.webp)
Oogenesis [PDF]
Oogenesis Dr Navneet Kumar Professor (Anatomy) K.G.M.U Dr NavneetKumar Professor Anatomy KGMU Lko Oogenesis • Development of ovum (oogenesis) • Maturation of follicle • Fate of ovum and follicle Dr NavneetKumar Professor Anatomy KGMU Lko Dr NavneetKumar Professor Anatomy KGMU Lko Oogenesis • Site – ovary • Duration – 7th week of embryo –primordial germ cells • -3rd month of fetus –oogonium • - two million primary oocyte • -7th month of fetus primary oocyte +primary follicle • - at birth primary oocyte with prophase of • 1st meiotic division • - 40 thousand primary oocyte in adult ovary • - 500 primary oocyte attain maturity • - oogenesis completed after fertilization Dr Navneet Kumar Dr NavneetKumar Professor Professor (Anatomy) Anatomy KGMU Lko K.G.M.U Development of ovum Oogonium(44XX) -In fetal ovary Primary oocyte (44XX) arrest till puberty in prophase of 1st phase meiotic division Secondary oocyte(22X)+Polar body(22X) 1st phase meiotic division completed at ovulation &enter in 2nd phase Ovum(22X)+polarbody(22X) After fertilization Dr NavneetKumar Professor Anatomy KGMU Lko Dr NavneetKumar Professor Anatomy KGMU Lko Dr Navneet Kumar Dr ProfessorNavneetKumar (Anatomy) Professor K.G.M.UAnatomy KGMU Lko Dr NavneetKumar Professor Anatomy KGMU Lko Maturation of follicle Dr NavneetKumar Professor Anatomy KGMU Lko Maturation of follicle Primordial follicle -Follicular cells Primary follicle -Zona pallucida -Granulosa cells Secondary follicle Antrum developed Ovarian /Graafian follicle - Theca interna &externa -Membrana granulosa -Antrial -

Pip Pip Cheerio! a Baby Chick Is Hatching Ever Wonder How a Baby Bird Gets out of Its Egg? It Has Got to Require Some Hard Work for Such a Tiny Little Animal
Pip Pip Cheerio! A Baby Chick is Hatching Ever wonder how a baby bird gets out of its egg? It has got to require some hard work for such a tiny little animal. Luckily, chicks are equipped with a handy dandy tool built into their beaks called an egg tooth. An egg tooth grows at the very tip of the beak. They are very small and sharp enough to help break through the shell of the egg. This is called pipping. Learn what it’s like to pip through an eggshell with this fun art activity. You will need: ● Two pieces of paper, any color will work, but white is the easiest to decorate! ● Markers, crayons, or colored pencils ● Scissors ● Glue (a glue stick will work well!) or double sided tape ● Optional: tweezers, spoon, fork, or other tools. Instructions: Step 1: Draw out an egg shape on the paper Step 2: Cut out two egg shaped pieces of paper about the same size *Helpful note: you can make them the same size by putting the two pieces of paper together and cutting them out at the same time Step 3: Decorate one side of the egg to make your chick! Get creative with your decorations but centering your chick’s face will give you the best results. The other piece will be the eggshell, which you can also decorate! *Helpful note: You will be ripping this paper later, so keep the decorations to the outside edges. Step 4: Take your chick paper and glue all around its edge; if you are using double-sided tape, tape around the edges. -

EM083E.Pdf (1.018Mb)
Poultry Leader Guide EM083E Level 3 4-H Poultry Leader Notebook Level III Poultry Dating Game ................................................................................3 Reproduction and Fertilization of Poultry ................................................7 Proper Handling of Hatching Eggs .........................................................15 Embryonic Mortality ...............................................................................19 General Hatchery Management Practices ...............................................21 The Chicks Are Here ...............................................................................27 Basic Nutritional Needs of Poultry .........................................................33 Controlling Body Weight of Replacement Birds ....................................37 The Comforts of Home ...........................................................................41 Light Sensitivity in Chickens .................................................................45 Managing a Small Laying Flock .............................................................51 Culling the Layer Flock ..........................................................................59 Adaptations for Flight .............................................................................63 Flight Prevention .....................................................................................67 Eggs: Normal and Irregular ....................................................................71 Marketing Eggs and Poultry Products -

Infertility Investigations for Women
Infertility investigations for women Brooke Building Gynaecology Department 0161 206 5224 © G21031001W. Design Services, Salford Royal NHS Foundation Trust, All Rights Reserved 2021. Document for issue as handout. Unique Identifier: SURG08(21). Review date: May 2023. This booklet is aimed for women undergoing fertility LH (Luteinising Hormone) Progesterone investigations. Its’ aim is to Oligomenorrhoea - When the provide you with some useful periods are occurring three In women, luteinising hormone Progesterone is a female information regarding your or four times a year (LH) is linked to ovarian hormone produced by the hormone production and egg ovaries after ovulation. It investigations. Irregular cycle - Periods that maturation. LH is used to causes the endometrial lining vary in length We hope you !nd this booklet measure a woman’s ovarian of the uterus to get thicker, helpful. The following blood tests are reserve (egg supply). making it receptive for a used to investigate whether You will be advised to have some It causes the follicles to grow, fertilised egg. ovulation (production of an egg) or all of the following tests: mature and release the eggs Progesterone levels increase is occurring each month and also for fertilisation. It reaches its after ovulation, reaching a to help determine which fertility Hormone blood tests highest level (the LH surge) in maximum level seven days treatments to offer. Follicular bloods tests the middle of the menstrual before the start of the next cycle 48 hours prior to ovulation period. The progesterone test is These routine blood tests are FSH (Follicle Stimulating i.e. days 12-14 of a 28 day cycle. -

Identifying Lethal Temperatures Targeting Immature Life Stage Control of Spotted Wing Drosophila
AGRICULTURAL RESEARCH FOUNDATION FINAL REPORT FUNDING CYCLE 2015 – 2017 TITLE: Identifying Lethal Temperatures Targeting Immature Life Stage Control of Spotted Wing Drosophila RESEARCH LEADER: Vaughn Walton COOPERATORS: Daniel Dalton, Riki York SUMMARY: Survival of spotted wing drosophila (D. suzukii, SWD) larvae to adulthood was evaluated after exposure to temperatures ranging 29-49°C in a laboratory thermal gradient bench for 60 minutes. Vials filled with pre-made fly diet up to approximately 1.5 cm from the bottom were used as oviposition media inside the gradient wells. Forty-three vials were prepared each day for 4 consecutive days. Five adult females were placed in each of 36 vials for 24 hours for oviposition to occur, seven vials were held as controls without flies. Upon removal, adult flies were placed in ethanol. Four ages were tested between 1-4-day old egg/larvae (i.e. 1 day old, 2 days old, 3 days old, and four days old). Sample vials were examined for oviposition success. Immature flies exposed temperatures of 32-35°C had the highest survival to adulthood. Above 38°C survival to adulthood with no survival at 41°C and 49°C, and a few surviving to adulthood under 45°C. An additional trial was conducted to evaluate the potential for thermal conditioning to improve larval survival at higher temperatures. For this study, four-day-old larvae were subject to 30, 60, and 90 minutes at 35°C, then allowed to cool to room temperature (22°C) for 60 minutes prior to placement in the thermal bench. These trials were used to determine how heat may reduce successful development to adulthood, leading to better management practices in the field. -
Understanding Your Menstrual Cycle If You're Trying to Conceive
IS MY PERIOD NORMAL? Understanding Your Menstrual Cycle If You’re Trying to Conceive More than 70% 11% 95% of women have or more of of U.S. women start irregular menstrual American women their periods by cycles as menopause suffer from age 16. approaches. endometriosis.1 10% 12% of U.S. women are of women have affected by PCOS trouble getting or (polycystic ovary staying pregnant.3 syndrome).2 Fortunately, your menstrual cycle can tell you a lot about your fertility if you know what to look for. TYPES OF MENSTRUAL CYCLES Only 15% of About Normal = women have 30% of women are fertile only during 21 to 35 days the “perfect” the “normal” fertility 28-day cycle. window—between days 10 and 17 of the menstrual cycle. Day 1 Period starts (aka menses) 27 28 1 2 26 3 25 4 24 5 Day 15-28 23 6 Day 2-14 Luteal phase; Follicular phase; progesterone** 22 WHAT’S NORMAL? 7 FSH released, (follicle- uterine lining 21 8 stimulating matures Give or take a few days, hormone) and a normal cycle looks like this: estrogen released, 20 9 ovulation* begins 19 10 18 11 17 12 16 15 14 13 *ovulation: the process of an ovum (egg) being released from the ovary; occurs 10-14 days before menses. **progesterone: a steroid hormone that tells the uterus to prepare for pregnancy At least 30% of women have an “irregular” cycle either short, long or inconsistent. Short = Long = < 21 days > 35 days May be a sign of: May be a sign of: Hormonal imbalance Hormonal imbalance Ovaries with fewer eggs Lack of ovulation Approach of menopause Other fertility issues Reduced fertility4 Increased risk of miscarriage SIGNS TO WATCH FOR Your menstrual cycle provides valuable clues about your body’s reproductive health. -
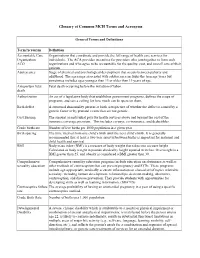
Glossary of Common MCH Terms and Acronyms
Glossary of Common MCH Terms and Acronyms General Terms and Definitions Term/Acronym Definition Accountable Care Organizations that coordinate and provide the full range of health care services for Organization individuals. The ACA provides incentives for providers who join together to form such ACO organizations and who agree to be accountable for the quality, cost, and overall care of their patients. Adolescence Stage of physical and psychological development that occurs between puberty and adulthood. The age range associated with adolescence includes the teen age years but sometimes includes ages younger than 13 or older than 19 years of age. Antepartum fetal Fetal death occurring before the initiation of labor. death Authorization An act of a legislative body that establishes government programs, defines the scope of programs, and sets a ceiling for how much can be spent on them. Birth defect A structural abnormality present at birth, irrespective of whether the defect is caused by a genetic factor or by prenatal events that are not genetic. Cost Sharing The amount an individual pays for health services above and beyond the cost of the insurance coverage premium. This includes co-pays, co-insurance, and deductibles. Crude birth rate Number of live births per 1000 population in a given year. Birth spacing The time interval from one child’s birth until the next child’s birth. It is generally recommended that at least a two-year interval between births is important for maternal and child health and survival. BMI Body mass index (BMI) is a measure of body weight that takes into account height.