A.4. Families of Sclaroldea
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
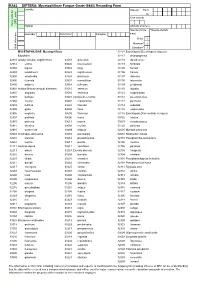
Ra82 Diptera
RA82 DIPTERA: Mycetophilinae Fungus Gnats (6480) Recording Form Locality Date(s) from: to: Vice county GPS users Grey cells for Habitat Altitude (metres) Source (circle *Source details Recorder Determiner Compiler one) Field 1 Museum* 2 Grid reference Literature* 3 MYCETOPHILIDAE: Mycetophilinae 33101 Exechiopsis (Exechiopsis) clypeata Exechiini 33117 dryaspagensis 32801 Allodia (Allodia) anglofennica 33519 griseolum33118 dumitrescae 32912 embla 33526 intermedium33119 fimbriata 32802 lugens 33522 kingi33120 furcata 32803 lundstroemi 33523 nigrofuscum33106 hammi 32804 ornaticollis 33524 proximum33107 indecisa 32806 truncata 33527 rosmellitum33108 intersecta 32805 zaitzevi 33514 ruficorne33109 jenkinsoni 32901 Allodia (Brachycampta) alternans 33515 serenum33110 ligulata 32910 angulata 33516 sericoma33121 magnicauda 32903 barbata 33601 Cordyla brevicornis33112 pseudindecisa 32904 czernyi 33602 crassicornis33113 pulchella 32909 foliifera 33603 fasciata33114 subulata 32905 grata 33604 fissa33115 unguiculata 32906 neglecta 33605 flaviceps33116 Exechiopsis (Xenexechia) crucigera 32907 pistillata 33606 fusca33002 leptura 32915 protenta 33613 insons33003 membranacea 32914 silvatica 33608 murina33122 pollicata 32916 westerholti 33609 nitidula32605 Myrosia maculosa 32602 Allodiopsis domestica 33610 parvipalpis32601 Notolopha cristata 32610 korolevi 33614 pseudomurina32701 Pseudexechia aurivernica 32607 rustica 33611 pusilla32706 monica 32212 Anatella alpina 33612 semiflava32705 parallela 32213 ankeli 33201 Exechia bicincta32703 trisignata 32216 bremia -

Recent Noteworthy Findings of Fungus Gnats from Finland and Northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae)
Biodiversity Data Journal 2: e1068 doi: 10.3897/BDJ.2.e1068 Taxonomic paper Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae) Jevgeni Jakovlev†, Jukka Salmela ‡,§, Alexei Polevoi|, Jouni Penttinen ¶, Noora-Annukka Vartija# † Finnish Environment Insitutute, Helsinki, Finland ‡ Metsähallitus (Natural Heritage Services), Rovaniemi, Finland § Zoological Museum, University of Turku, Turku, Finland | Forest Research Institute KarRC RAS, Petrozavodsk, Russia ¶ Metsähallitus (Natural Heritage Services), Jyväskylä, Finland # Toivakka, Myllyntie, Finland Corresponding author: Jukka Salmela ([email protected]) Academic editor: Vladimir Blagoderov Received: 10 Feb 2014 | Accepted: 01 Apr 2014 | Published: 02 Apr 2014 Citation: Jakovlev J, Salmela J, Polevoi A, Penttinen J, Vartija N (2014) Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae). Biodiversity Data Journal 2: e1068. doi: 10.3897/BDJ.2.e1068 Abstract New faunistic data on fungus gnats (Diptera: Sciaroidea excluding Sciaridae) from Finland and NW Russia (Karelia and Murmansk Region) are presented. A total of 64 and 34 species are reported for the first time form Finland and Russian Karelia, respectively. Nine of the species are also new for the European fauna: Mycomya shewelli Väisänen, 1984,M. thula Väisänen, 1984, Acnemia trifida Zaitzev, 1982, Coelosia gracilis Johannsen, 1912, Orfelia krivosheinae Zaitzev, 1994, Mycetophila biformis Maximova, 2002, M. monstera Maximova, 2002, M. uschaica Subbotina & Maximova, 2011 and Trichonta palustris Maximova, 2002. Keywords Sciaroidea, Fennoscandia, faunistics © Jakovlev J et al. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. -

Lepidoptera: Geometridae, Geometrinae) SHILAP Revista De Lepidopterología, Vol
SHILAP Revista de Lepidopterología ISSN: 0300-5267 [email protected] Sociedad Hispano-Luso-Americana de Lepidopterología España Viidalepp, J.; Lindt, A. A new Rhanidopsis West, 1930 from the Malay Peninsula (Lepidoptera: Geometridae, Geometrinae) SHILAP Revista de Lepidopterología, vol. 38, núm. 149, marzo, 2010, pp. 97-106 Sociedad Hispano-Luso-Americana de Lepidopterología Madrid, España Available in: http://www.redalyc.org/articulo.oa?id=45514996008 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative 97-106 A new Rhanidopsis West, 7/3/10 16:45 Página 97 SHILAP Revta. lepid., 38 (149), marzo 2010: 97-106 CODEN: SRLPEF ISSN:0300-5267 A new Rhanidopsis West, 1930 from the Malay Peninsula (Lepidoptera: Geometridae, Geometrinae) J. Viidalepp & A. Lindt Abstract A new moth species from the oriental geometrid genus Rhanidopsis West, 1930 is described: R. kogeri Viidalepp & Lindt, sp. n., from Cameron Highlands, Peninsular Malaysia is added to the distribution area of the genus, formerly known from the Philippines and Borneo. Phenotypical similarity of the genus with Afrotropical Dargeia Herbulot, 1977 and Neotropical Pyrochlora Warren, 1895 is discussed. Some plesiomorphic characters of Neotropical and Indoaustralian genera of Nemoriini are compared. KEY WORDS: Lepidoptera, Geometridae, Geometrinae, Pyrochlora, Dargeia, Rhanidopsis kogeri, new species, Malay Peninsula. Un nuevo Rhanidopsis West, 1930 de Malasia (Lepidoptera: Geometridae, Geometrinae) Resumen Se describe una nueva especie del género geométrido oriental Rhanidopsis West, 1930: R. kogeri Viidalepp & Lindt, sp. -

Zootaxa, the Fungus Gnats (Diptera: Bolitophilidae
Zootaxa 2318: 450–506 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) The fungus gnats (Diptera: Bolitophilidae, Keroplatidae, Mycetophilidae) of Sardinia, with description of six new species* PETER J. CHANDLER 606B Berryfield Lane, Melksham, Wilts SN12 6EL, United Kingdom. E-mail: [email protected] *In: Cerretti, P., Mason, F., Minelli, A., Nardi, G. & Whitmore, D. (Eds), Research on the Terrestrial Arthropods of Sardinia (Italy). Zootaxa, 2318, 1–602. Table of contents Abstract . .450 Introduction . .451 Study area . .452 Material and methods . .452 Abbreviations . .453 Sampling sites . .454 Faunistic list . .456 Bolitophilidae . .456 Keroplatidae, Keroplatinae, Keroplatini . .456 Orfeliini . .457 Macrocerinae . .462 Mycetophilidae, Gnoristinae . .465 Leiinae . .469 Mycetophilinae, Exechiini . .472 Mycetophilini . .480 Mycomyinae . .492 Sciophilinae . .495 Discussion . .500 Acknowledgements . .501 References . .502 Abstract The fungus gnat fauna of Sardinia is reviewed and data presented for all species recorded. Altogether one species of Bolitophilidae, 16 species of Keroplatidae and 105 species of Mycetophilidae are recognised as occurring in Sardinia. As the bolitophilid and two of the mycetophilid species are represented only by females and are not determined to species level, the total confirmed Sardinian list stands at 119 species. Four species of Keroplatidae and 19 species of Mycetophilidae are new to the total Italian fauna, whereas three species of Keroplatidae and 32 species of Mycetophilidae are newly recorded for the island of Sardinia. Six species are described as new to science: two Keroplatidae (Urytalpa juliae sp. nov., Macrocera nuragica sp. nov.) and four Mycetophilidae (Boletina ichnusa sp. nov., Trichonta sandalyon sp. -

Diptera) Diversity in a Patch of Costa Rican Cloud Forest: Why Inventory Is a Vital Science
Zootaxa 4402 (1): 053–090 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2018 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4402.1.3 http://zoobank.org/urn:lsid:zoobank.org:pub:C2FAF702-664B-4E21-B4AE-404F85210A12 Remarkable fly (Diptera) diversity in a patch of Costa Rican cloud forest: Why inventory is a vital science ART BORKENT1, BRIAN V. BROWN2, PETER H. ADLER3, DALTON DE SOUZA AMORIM4, KEVIN BARBER5, DANIEL BICKEL6, STEPHANIE BOUCHER7, SCOTT E. BROOKS8, JOHN BURGER9, Z.L. BURINGTON10, RENATO S. CAPELLARI11, DANIEL N.R. COSTA12, JEFFREY M. CUMMING8, GREG CURLER13, CARL W. DICK14, J.H. EPLER15, ERIC FISHER16, STEPHEN D. GAIMARI17, JON GELHAUS18, DAVID A. GRIMALDI19, JOHN HASH20, MARTIN HAUSER17, HEIKKI HIPPA21, SERGIO IBÁÑEZ- BERNAL22, MATHIAS JASCHHOF23, ELENA P. KAMENEVA24, PETER H. KERR17, VALERY KORNEYEV24, CHESLAVO A. KORYTKOWSKI†, GIAR-ANN KUNG2, GUNNAR MIKALSEN KVIFTE25, OWEN LONSDALE26, STEPHEN A. MARSHALL27, WAYNE N. MATHIS28, VERNER MICHELSEN29, STEFAN NAGLIS30, ALLEN L. NORRBOM31, STEVEN PAIERO27, THOMAS PAPE32, ALESSANDRE PEREIRA- COLAVITE33, MARC POLLET34, SABRINA ROCHEFORT7, ALESSANDRA RUNG17, JUSTIN B. RUNYON35, JADE SAVAGE36, VERA C. SILVA37, BRADLEY J. SINCLAIR38, JEFFREY H. SKEVINGTON8, JOHN O. STIREMAN III10, JOHN SWANN39, PEKKA VILKAMAA40, TERRY WHEELER††, TERRY WHITWORTH41, MARIA WONG2, D. MONTY WOOD8, NORMAN WOODLEY42, TIFFANY YAU27, THOMAS J. ZAVORTINK43 & MANUEL A. ZUMBADO44 †—deceased. Formerly with the Universidad de Panama ††—deceased. Formerly at McGill University, Canada 1. Research Associate, Royal British Columbia Museum and the American Museum of Natural History, 691-8th Ave. SE, Salmon Arm, BC, V1E 2C2, Canada. Email: [email protected] 2. -

Diptera; Diadocidiidae and Mycetophilidae)
Fungus gnats from Jostedalen, West Norway (Diptera; Diadocidiidae and Mycetophilidae) GEIR E. E. SBLI Snli, G. E. E. 1994. Fungus gnats from Jostedalen, West Norway (Diptera: Diadocidi- idae and Mycetophilidae). Fauna norv. Ser. B 41: 1-12. During a study of terrestrial invertebrates in Jostedalen in 1988, more than 3.000 specimens of fungus gnats were caught. 214 species were recognized, belonging to the families Diadocidiidae and Mycetophilidae. The number of species in Jostedalen is exceptionally high when compared to number of species recorded from other local areas in Europe. The genus Drepanocercus (Vockeroth, 1980) is recorded for the first time from the Palaearctic region. Other rare species are Mycomya simulans Vaisanen, 1984, Acnemia falcata Zaitzev, 1982, Zygom.via pseudohumeralis Caspers, 1980, Anatella aquila Zaitsev, 1989. A. fungina Plassmann, 1984, Exechia subfrigida Las- tovka & Matila, 1974. Exechiopsis dryaspagensis Chandler. 1977 and E. pseudopul- chella (Lundstrom, 1909). Twenty species could not be identified, half of which undoubtly represent undescribed species. The fauna of Norwegian fungus gnats is poorly documented, and most species recorded here are new to Norway. According to the present knowledge on the distribution of fungus gnats, the fauna in Jostedalen seems to have an affinity to the central/eastern Palaearctic fauna, and has more species in common with the Finnish fauna than with the British. Geir E. E. Snli, Museum of Zoology, University of Bergen, Musiplass 3, N-5007 Bergen, Norway. INTRODUCTION The river Jostedola has its origin in the gla- dae, Sciardae and Mycetophilidae. Fungus cier Jostedalsbreen, the largest ice cap on the gnats are distributed all over the world, but European mainland, and runs through the their taxonomy, biology and biogeography valley Jostedalen. -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

Zootaxa, Diptera, Mycetophilidae
Zootaxa 856: 1–35 (2005) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA 856 Copyright © 2005 Magnolia Press ISSN 1175-5334 (online edition) A review of fungus gnats in the tribe Exechiini (Diptera, Myceto- philidae) from the J. W. Zetterstedt collection at the Museum of Zoology in Lund, Sweden JOSTEIN KJÆRANDSEN Museum of Zoology, Lund University, Helgonavägen 3, S-223 62 Lund, Sweden. ([email protected]) Table of contents Abstract . 1 Introduction . 2 Material and methods . 4 Result . 6 List of species . 12 Acknowledgements . 33 References . 33 Abstract The collections of fungus gnats by Johan Wilhelm Zetterstedt (1785–1874), lodged in the Museum of Zoology in Lund, Sweden, are examined for all species belonging in the tribe Exechiini Edwards. The majority of the material was collected in Fennoscandia, mainly in Sweden, in the first half of the 19th century. Altogether 37 species of the tribe Exechiini could be safely identified. Three additional species are strongly indicated to be present in the collections, but could not be identified with certainty, viz. Allodia (Brachycampta) alternans (Zetterstedt, 1838), Cordyla murina Winnertz, 1863 and Stigmatomeria crassicornis (Stannius, 1831). Some of Zetterstedt's types have been erroneously synonymized and misinterpreted in modern literature. Hence, illustra- tions of terminalia are presented for all recognizable Exechiini types described by Zetterstedt. In order to preserve nomenclatural stability a lectotype is selected for Brevicornu griseolum (Zetter- stedt, 1852) sensu auctore nec Edwards, and a neotype is selected for Allodia (Brachycampta) alternans (Zetterstedt, 1838). Two species names are reinstated, viz. Brevicornu canescens (Zetter- stedt, 1852) sp. -

Fungus Gnats
Евразиатский энтомол. журнал 16(2): 119–128 © EUROASIAN ENTOMOLOGICAL JOURNAL, 2017 Fungus gnats (Diptera: Bolitophilidae, Diadocidiidae, Keroplatidae, Mycetophilidae) of the lower course of Anadyr River, Chukotskii Autonomnyi Okrug, Russia Ãðèáíûå êîìàðû (Diptera, Syrphidae) íèçîâèé ðåêè Àíàäûðü (×óêîòñêèé àâòîíîìíûé îêðóã, Ðîññèÿ) A.V. Polevoi*, A.V. Barkalov** À.Â. Ïîëåâîé*, À.Â. Áàðêàëîâ** * Forest Research Institute, Karelian Research Centre of the Russian Academy of Sciences, Pushkinskaya Str. 11, Petrozavodsk 185910 Russia. E-mail: [email protected]. * Институт леса КарНЦ РАН, ул. Пушкинская 11, Петрозаводск, 185910, Россия. E-mail: [email protected] ** Institute of Systematics and Ecology of Animals, Russian Academy of Sciences, Siberian Branch, Frunze Str. 11, Novosibirsk 630091 Russia. E-mail: [email protected]. ** Институт систематики и экологии животных СО РАН, ул. Фрунзе 11, Новосибирск 630091 Россия. Key words: fauna, fungus gnats, Anadyr River, Chukotka. Ключевые слова: фауна, грибные комары, река Анадырь, Чукотка. Abstract. The first data on the Fungus gnats fauna of Diadocidiidae, Ditomyiidae, Keroplatidae and Myceto- Chukotka are presented. 170 species belonging to the fami- philidae, belonging to the superfamily Sciaroidea (Diptera, lies Bolitophilidae, Diadocidiidae, Keroplatidae and Myce- Nematocera, Bibionomorpha). This is highly diverse group tophilidae were reported during two field seasons in 2013 with estimated number of species around 4500 in the and 2014 in the lower course of the Anadyr River. Eight world fauna and more than 1450 in Palaearctic [Søli et al., species are reported from Russia for the first time, two species are new for the Palaearctic and 27 species were 2000]. In the latter region, they seem to display an in- previously unknown in the eastern part of the Palaearctic; creasing diversity towards the North, with most species- 28 species are most probably undescribed taxa. -

Kjaerandsen Sciaroidea WIP.Pdf
Species recognition trade-off between structural wing colours and terminalia in fungus gnats ? J. Kjaerandsen Museum of Zoology Lund University Sweden Structural colours in flies Reflective scales in Diptera – Mosquitoes: Toxorhynchites manicatus (Japan) Reflective body scales in fungus gnats – only in the genus Allactoneura ? Hymenoptera: Eulophidae PhD student Ekaterina Shevtsova Wings imbedded in a medium or studied on a white background will not display their structural colours Slide with wings embedded in Canada balsam Dry specimens studied on a pure white background Mycetophilidae: Rymosia fasciata Keroplatidae: Proceroplatus scalprifera WIPs — Wing Interference Patterns i for interference Bolitophila occlusa Hybotidae: Ocydromia glabricula Cordyla sp. (California) Exechia nugatoria Photo: “Klaas” at Diptera.info, 2008 (= nigroscutellata) (California) Photos: Peter Kerr, 2008 My photo of the same species’ WIP Photos of structural wing colours on internet WIPs — Wing Interference Patterns i for interference • — Genetics of pigment patterns • — Thin Film Interference • — Newton Scale Metering • — Exechiini • — Lygistorrhinidae • — Keroplatidae • — The trade-off Pigmentation in brown, yellow and black: Spatiotemporally regulated by yellow and ebony MELANINS Leia Proceroplatus (Japan) (New Caledonia) Scientists unlock mystery of animal colour patterns Genetic April 22 control of pigment 2010 patterns T. Werner, S. Koshikawa, T. M. Williams, S. B. Carroll, Nature 464, 1143 (2010) Pigments are only a part of the ”mystery of wing colour -

Smithsonian Miscellaneous Collections Volume 117, Number 3
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 117, NUMBER 3 RELATIONSHIPS OF CERTAIN GENERA OF FUNGUS GNATS OF THE FAMILY MYCETOPHILIDAE BY F. R. SHAW AND M. M. SHAW Amherst, Mass. (Publication 4053) CITY OF WASHINGTON PUBLISHED BY THE SMITHSONIAN INSTITUTION DECEMBER 27, 1951 Z^t £or5 (g»af(tmore (pv&ee BALTIMOEB, MD., 0. B. A. RELATIONSHIPS OF CERTAIN GENERA OF FUNGUS GNATS OF THE FAMILY MYCETOPHILIDAE By F. R. SHAW and M. M. SHAW i Amherst, Mass. The present study represents a continuation of a preliminary in- vestigation of the possible value of thoracic sclerites in determining the relationships of certain insects. Dr. G. C. Crampton was the first to demonstrate the use of these sclerites as a means of determin- ing the systematic position of insects. In 1925, he published a clas- sical study of the comparative morphology of the thorax of nontipu- loid Nematocera. In 1948 Shaw presented a paper in which he indi- cated the value of thoracic sclerites as an aid in determining the phy- logeny of the Mycetophilidae. Although the number of genera he studied was admittedly small, principles were developed that have been of value in distinguishing the phylogenetic relationships of cer- tain genera. Edwards (1925) was the first to indicate that the structure of thoracic sclerites might be of value in determining generic characters in this group. In his monograph of the British fungus gnats he noted that in certain genera the sclerites differed in form and that such dif- ferences might be of value in separating groups of these insects. 1 We wish to express our thanks to the Society of Sigma Xi for a grant-in- aid that made possible the preparation of the illustrations for this paper. -

Insect Egg Size and Shape Evolve with Ecology but Not Developmental Rate Samuel H
ARTICLE https://doi.org/10.1038/s41586-019-1302-4 Insect egg size and shape evolve with ecology but not developmental rate Samuel H. Church1,4*, Seth Donoughe1,3,4, Bruno A. S. de Medeiros1 & Cassandra G. Extavour1,2* Over the course of evolution, organism size has diversified markedly. Changes in size are thought to have occurred because of developmental, morphological and/or ecological pressures. To perform phylogenetic tests of the potential effects of these pressures, here we generated a dataset of more than ten thousand descriptions of insect eggs, and combined these with genetic and life-history datasets. We show that, across eight orders of magnitude of variation in egg volume, the relationship between size and shape itself evolves, such that previously predicted global patterns of scaling do not adequately explain the diversity in egg shapes. We show that egg size is not correlated with developmental rate and that, for many insects, egg size is not correlated with adult body size. Instead, we find that the evolution of parasitoidism and aquatic oviposition help to explain the diversification in the size and shape of insect eggs. Our study suggests that where eggs are laid, rather than universal allometric constants, underlies the evolution of insect egg size and shape. Size is a fundamental factor in many biological processes. The size of an 526 families and every currently described extant hexapod order24 organism may affect interactions both with other organisms and with (Fig. 1a and Supplementary Fig. 1). We combined this dataset with the environment1,2, it scales with features of morphology and physi- backbone hexapod phylogenies25,26 that we enriched to include taxa ology3, and larger animals often have higher fitness4.