Smithsonian Miscellaneous Collections Volume 117, Number 3
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diptera, Nematocera) from the Mediterranean Region*)
©Zoologische Staatssammlung München;download: http://www.biodiversitylibrary.org/; www.biologiezentrum.at SPIXIANA 14 321-338 München, 3 1 . Oktober 1 991 ISSN0341-8391 New and remarkable species of Mycetophiloidea (Diptera, Nematocera) from the Mediterranean region*) By Norbert Caspers Caspers, N. (1991): New and remarkable species of Mycetophiloidea (Diptera, Nematocera) from the Mediterranean region. - Spixiana 14/3: 321-338. Descriptions of adult males of six neu species of the superfamily Mycetophiloidea (Diptera, Nematocera) from the Mediterranean region are presented: Orfelia persimi- lis, spec. nov. (family Keroplatidae) from Sardinia (Italy); Ectrepesthoneura chand- leri, spec. nov. from Crete (Greece), Leia umbrosa, spec. nov. from Corsica (France), Anatella longiflagellata, spec. nov. and Phronia incisa spec. nov. from Turkey, Sceptonia curvisetosa, spec. nov. from Greece, last five belonging to the family Mycetophilidae sensu Malloch (1917). In addition, new findings of two remarkable and hitherto rarely found species of Mycetophiloidea (Sciopbila kasbmirensis Zaitzev, Phronia tyrrhenica Edwards) are presented. Prof. Dr. Norbert Caspers, St.-Maternus-Eck 14 a, W-5067 Bechen, Germanv Introduction Since the early seventies Dr. H. Malicky (Biological Station Lunz, Austria) has been performing many entomological field trips to Southern Europe, with the aim to clear up the systematics and the biogeographic distribution of the caddis flies (Trichoptera) of that previously rather neglected faunal region. Besides Trichoptera he also collected a comprehensive material of many other aquatic insect groups, which meanwhile has been worked upon and has been published bv different authors. Among the "non-target-groups" of this study the portion of the fungus gnats (Mycetophiloidea) was relatively low as compared to all aquatic insect groups. -

The 2014 Golden Gate National Parks Bioblitz - Data Management and the Event Species List Achieving a Quality Dataset from a Large Scale Event
National Park Service U.S. Department of the Interior Natural Resource Stewardship and Science The 2014 Golden Gate National Parks BioBlitz - Data Management and the Event Species List Achieving a Quality Dataset from a Large Scale Event Natural Resource Report NPS/GOGA/NRR—2016/1147 ON THIS PAGE Photograph of BioBlitz participants conducting data entry into iNaturalist. Photograph courtesy of the National Park Service. ON THE COVER Photograph of BioBlitz participants collecting aquatic species data in the Presidio of San Francisco. Photograph courtesy of National Park Service. The 2014 Golden Gate National Parks BioBlitz - Data Management and the Event Species List Achieving a Quality Dataset from a Large Scale Event Natural Resource Report NPS/GOGA/NRR—2016/1147 Elizabeth Edson1, Michelle O’Herron1, Alison Forrestel2, Daniel George3 1Golden Gate Parks Conservancy Building 201 Fort Mason San Francisco, CA 94129 2National Park Service. Golden Gate National Recreation Area Fort Cronkhite, Bldg. 1061 Sausalito, CA 94965 3National Park Service. San Francisco Bay Area Network Inventory & Monitoring Program Manager Fort Cronkhite, Bldg. 1063 Sausalito, CA 94965 March 2016 U.S. Department of the Interior National Park Service Natural Resource Stewardship and Science Fort Collins, Colorado The National Park Service, Natural Resource Stewardship and Science office in Fort Collins, Colorado, publishes a range of reports that address natural resource topics. These reports are of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Report Series is used to disseminate comprehensive information and analysis about natural resources and related topics concerning lands managed by the National Park Service. -
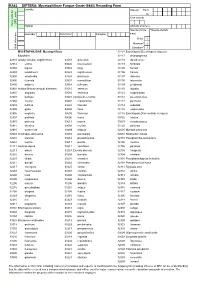
Ra82 Diptera
RA82 DIPTERA: Mycetophilinae Fungus Gnats (6480) Recording Form Locality Date(s) from: to: Vice county GPS users Grey cells for Habitat Altitude (metres) Source (circle *Source details Recorder Determiner Compiler one) Field 1 Museum* 2 Grid reference Literature* 3 MYCETOPHILIDAE: Mycetophilinae 33101 Exechiopsis (Exechiopsis) clypeata Exechiini 33117 dryaspagensis 32801 Allodia (Allodia) anglofennica 33519 griseolum33118 dumitrescae 32912 embla 33526 intermedium33119 fimbriata 32802 lugens 33522 kingi33120 furcata 32803 lundstroemi 33523 nigrofuscum33106 hammi 32804 ornaticollis 33524 proximum33107 indecisa 32806 truncata 33527 rosmellitum33108 intersecta 32805 zaitzevi 33514 ruficorne33109 jenkinsoni 32901 Allodia (Brachycampta) alternans 33515 serenum33110 ligulata 32910 angulata 33516 sericoma33121 magnicauda 32903 barbata 33601 Cordyla brevicornis33112 pseudindecisa 32904 czernyi 33602 crassicornis33113 pulchella 32909 foliifera 33603 fasciata33114 subulata 32905 grata 33604 fissa33115 unguiculata 32906 neglecta 33605 flaviceps33116 Exechiopsis (Xenexechia) crucigera 32907 pistillata 33606 fusca33002 leptura 32915 protenta 33613 insons33003 membranacea 32914 silvatica 33608 murina33122 pollicata 32916 westerholti 33609 nitidula32605 Myrosia maculosa 32602 Allodiopsis domestica 33610 parvipalpis32601 Notolopha cristata 32610 korolevi 33614 pseudomurina32701 Pseudexechia aurivernica 32607 rustica 33611 pusilla32706 monica 32212 Anatella alpina 33612 semiflava32705 parallela 32213 ankeli 33201 Exechia bicincta32703 trisignata 32216 bremia -

The Nestling Diet of Svalbard Snow Buntings Identified by DNA Metabarcoding
Faculty of Biosciences, Fisheries and Economics, Department of Arctic and Marine Biology The nestling diet of Svalbard snow buntings identified by DNA metabarcoding — Christian Stolz BIO-3950 Master thesis in Biology, Northern Populations and Ecosystems, May 2019 Faculty of Biosciences, Fisheries and Economics, Department of Arctic and Marine Biology The nestling diet of Svalbard snow buntings identified by DNA metabarcoding Christian Stolz, UiT The Arctic University of Norway, Tromsø, Norway and The University Centre in Svalbard (UNIS), Longyearbyen, Norway BIO-3950 Master Thesis in Biology, Northern Populations and Ecosystems, May 2018 Supervisors: Frode Fossøy, Norwegian Institute for Nature Research (NINA), Trondheim, Norway Øystein Varpe, The University Centre in Svalbard (UNIS), Longyearbyen, Norway Rolf Anker Ims, UiT The Arctic University of Norway, Tromsø, Norway i Abstract Tundra arthropods have considerable ecological importance as a food source for several bird species that are reproducing in the Arctic. The actual arthropod taxa comprising the chick diet are however rarely known, complicating assessments of ecological interactions. In this study, I identified the nestling diet of Svalbard snow bunting (Plectrophenax nivalis) for the first time. Faecal samples of snow bunting chicks were collected in Adventdalen, Svalbard in the breeding season 2018 and analysed via DNA metabarcoding. Simultaneously, the availability of prey arthropods was measured via pitfall trapping. The occurrence of 32 identified prey taxa in the nestling diet changed according to varying abundances and emergence patterns within the tun- dra arthropod community: Snow buntings provisioned their offspring mainly with the most abundant prey items which were in the early season different Chironomidae (Diptera) taxa and Scathophaga furcata (Diptera: Scathophagidae), followed by Spilogona dorsata (Diptera: Mus- cidae). -

Recent Noteworthy Findings of Fungus Gnats from Finland and Northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae)
Biodiversity Data Journal 2: e1068 doi: 10.3897/BDJ.2.e1068 Taxonomic paper Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae) Jevgeni Jakovlev†, Jukka Salmela ‡,§, Alexei Polevoi|, Jouni Penttinen ¶, Noora-Annukka Vartija# † Finnish Environment Insitutute, Helsinki, Finland ‡ Metsähallitus (Natural Heritage Services), Rovaniemi, Finland § Zoological Museum, University of Turku, Turku, Finland | Forest Research Institute KarRC RAS, Petrozavodsk, Russia ¶ Metsähallitus (Natural Heritage Services), Jyväskylä, Finland # Toivakka, Myllyntie, Finland Corresponding author: Jukka Salmela ([email protected]) Academic editor: Vladimir Blagoderov Received: 10 Feb 2014 | Accepted: 01 Apr 2014 | Published: 02 Apr 2014 Citation: Jakovlev J, Salmela J, Polevoi A, Penttinen J, Vartija N (2014) Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae). Biodiversity Data Journal 2: e1068. doi: 10.3897/BDJ.2.e1068 Abstract New faunistic data on fungus gnats (Diptera: Sciaroidea excluding Sciaridae) from Finland and NW Russia (Karelia and Murmansk Region) are presented. A total of 64 and 34 species are reported for the first time form Finland and Russian Karelia, respectively. Nine of the species are also new for the European fauna: Mycomya shewelli Väisänen, 1984,M. thula Väisänen, 1984, Acnemia trifida Zaitzev, 1982, Coelosia gracilis Johannsen, 1912, Orfelia krivosheinae Zaitzev, 1994, Mycetophila biformis Maximova, 2002, M. monstera Maximova, 2002, M. uschaica Subbotina & Maximova, 2011 and Trichonta palustris Maximova, 2002. Keywords Sciaroidea, Fennoscandia, faunistics © Jakovlev J et al. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. -

Zootaxa, the Fungus Gnats (Diptera: Bolitophilidae
Zootaxa 2318: 450–506 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) The fungus gnats (Diptera: Bolitophilidae, Keroplatidae, Mycetophilidae) of Sardinia, with description of six new species* PETER J. CHANDLER 606B Berryfield Lane, Melksham, Wilts SN12 6EL, United Kingdom. E-mail: [email protected] *In: Cerretti, P., Mason, F., Minelli, A., Nardi, G. & Whitmore, D. (Eds), Research on the Terrestrial Arthropods of Sardinia (Italy). Zootaxa, 2318, 1–602. Table of contents Abstract . .450 Introduction . .451 Study area . .452 Material and methods . .452 Abbreviations . .453 Sampling sites . .454 Faunistic list . .456 Bolitophilidae . .456 Keroplatidae, Keroplatinae, Keroplatini . .456 Orfeliini . .457 Macrocerinae . .462 Mycetophilidae, Gnoristinae . .465 Leiinae . .469 Mycetophilinae, Exechiini . .472 Mycetophilini . .480 Mycomyinae . .492 Sciophilinae . .495 Discussion . .500 Acknowledgements . .501 References . .502 Abstract The fungus gnat fauna of Sardinia is reviewed and data presented for all species recorded. Altogether one species of Bolitophilidae, 16 species of Keroplatidae and 105 species of Mycetophilidae are recognised as occurring in Sardinia. As the bolitophilid and two of the mycetophilid species are represented only by females and are not determined to species level, the total confirmed Sardinian list stands at 119 species. Four species of Keroplatidae and 19 species of Mycetophilidae are new to the total Italian fauna, whereas three species of Keroplatidae and 32 species of Mycetophilidae are newly recorded for the island of Sardinia. Six species are described as new to science: two Keroplatidae (Urytalpa juliae sp. nov., Macrocera nuragica sp. nov.) and four Mycetophilidae (Boletina ichnusa sp. nov., Trichonta sandalyon sp. -

Nomenclatural Studies Toward a World List of Diptera Genus-Group Names
Nomenclatural studies toward a world list of Diptera genus-group names. Part V Pierre-Justin-Marie Macquart Evenhuis, Neal L.; Pape, Thomas; Pont, Adrian C. DOI: 10.11646/zootaxa.4172.1.1 Publication date: 2016 Document version Publisher's PDF, also known as Version of record Document license: CC BY Citation for published version (APA): Evenhuis, N. L., Pape, T., & Pont, A. C. (2016). Nomenclatural studies toward a world list of Diptera genus- group names. Part V: Pierre-Justin-Marie Macquart. Magnolia Press. Zootaxa Vol. 4172 No. 1 https://doi.org/10.11646/zootaxa.4172.1.1 Download date: 02. Oct. 2021 Zootaxa 4172 (1): 001–211 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Monograph ZOOTAXA Copyright © 2016 Magnolia Press ISSN 1175-5334 (online edition) http://doi.org/10.11646/zootaxa.4172.1.1 http://zoobank.org/urn:lsid:zoobank.org:pub:22128906-32FA-4A80-85D6-10F114E81A7B ZOOTAXA 4172 Nomenclatural Studies Toward a World List of Diptera Genus-Group Names. Part V: Pierre-Justin-Marie Macquart NEAL L. EVENHUIS1, THOMAS PAPE2 & ADRIAN C. PONT3 1 J. Linsley Gressitt Center for Entomological Research, Bishop Museum, 1525 Bernice Street, Honolulu, Hawaii 96817-2704, USA. E-mail: [email protected] 2 Natural History Museum of Denmark, Universitetsparken 15, 2100 Copenhagen, Denmark. E-mail: [email protected] 3Oxford University Museum of Natural History, Parks Road, Oxford OX1 3PW, UK. E-mail: [email protected] Magnolia Press Auckland, New Zealand Accepted by D. Whitmore: 15 Aug. 2016; published: 30 Sept. 2016 Licensed under a Creative Commons Attribution License http://creativecommons.org/licenses/by/3.0 NEAL L. -

Supplementary Information for Evolution of Gene-Rich Germline Restricted
1 Supplementary Information for Evolution of gene-rich germline restricted 2 chromosomes in black-winged fungus gnats through introgression (Diptera: 3 Sciaridae) 4 Christina N. Hodson, Kamil S. Jaron, Susan Gerbi, Laura Ross 5 6 Supplementary Text 1: Detailed description of the chromosome inheritance system in 7 Bradysia coprophila. 8 9 The chromosome system in B. coprophila, and in sciarids generally, is unique in 10 several ways including chromosome transmission patterns, sex determination, and the 11 presence of GRCs (see Fig 1 for transmission patterns). All sciarids studied to date have a 12 system of reproduction known as paternal genome elimination, where males only transmit 13 maternally inherited chromosomes to offspring [1,2]. Paternal genome elimination has 14 evolved independently in at least seven arthropod lineages, including the related gall gnat 15 family Cecidomyiidae [3]. In all species with paternal genome elimination, meiosis occurs in 16 a Mendelian manner in females, but in males meiosis is aberrant. In male meiosis in 17 sciarids, there is a monopolar spindle in meiosis I. Maternally inherited chromosomes move 18 towards the monopolar spindle, while paternally derived chromosomes move away from it 19 and are discarded in a bud of cytoplasm [2]. Thus, only the maternal complement of 20 chromosomes is transmitted to the sperm. This phenomenon in B. coprophila was the first 21 example of “imprinting”, to our knowledge, by which the cell recognizes the maternal or 22 paternal origin of a chromosome [4]. Interestingly, the GRCs always segregate with the 23 maternal set of chromosomes. Therefore, all of the GRCs (typically two in B. -

Patterns of Animal Dispersal, Vicariance and Diversification in the Holarctic
Biological Journal of the Linnean Society (2001), 73: 345-390. With 15 figures doi:10.1006/bij1.2001.0542, available online at http;//www.idealibrary.comon IDE bl 0 c Patterns of animal dispersal, vicariance and diversification in the Holarctic ISABEL SANMARTIN1*, HENRIK ENGHOFF' and FREDRIK RONQUISTl 'Department of Systematic Zoology, Evolutionary Biology Centre, Uppsala University, Norbyvugen 180, SE-752 36 Uppsala, Sweden 2Zoologisk Museum, Uniuersitetsparken 15, DK-2100 Copenhagen, Denmark Received 23 October 2000; accepted for publication 25 March 2001 We analysed patterns of animal dispersal, vicariance and diversification in the Holarctic based on complete phylogenies of 57 extant non-marine taxa, together comprising 770 species, documenting biogeographic events from the Late Mesozoic to the present. Four major areas, each corresponding to a historically persistent landmass, were used in the analyses: eastern Nearctic (EN), western Nearctic (WN), eastern Palaeoarctic (EP) and western Palaeoarctic (WP). Parsimony-based tree fitting showed that there is no significantly supported general area cladogram for the dataset. Yet, distributions are strongly phylogenetically conserved, as revealed by dispersal- vicariance analysis (DIVA). DIVA-based permutation tests were used to pinpoint phylogenetically determined biogeographic patterns. Consistent with expectations, continental dispersals (WP-EP and WN-EN) are sig- nificantly more common than palaeocontinental dispersals (WN-EP and EN-WP), which in turn are more common than disjunct dispersals (EN-EP and WN-WP). There is significant dispersal asymmetry both within the Nearctic (WN+EN more common than EN+WN) and the Palaeoarctic (EP+WP more common than WP-tEP). Cross- Beringian faunal connections have traditionally been emphasized but are not more important than cross-Atlantic connections in our data set. -

Insect Egg Size and Shape Evolve with Ecology but Not Developmental Rate Samuel H
ARTICLE https://doi.org/10.1038/s41586-019-1302-4 Insect egg size and shape evolve with ecology but not developmental rate Samuel H. Church1,4*, Seth Donoughe1,3,4, Bruno A. S. de Medeiros1 & Cassandra G. Extavour1,2* Over the course of evolution, organism size has diversified markedly. Changes in size are thought to have occurred because of developmental, morphological and/or ecological pressures. To perform phylogenetic tests of the potential effects of these pressures, here we generated a dataset of more than ten thousand descriptions of insect eggs, and combined these with genetic and life-history datasets. We show that, across eight orders of magnitude of variation in egg volume, the relationship between size and shape itself evolves, such that previously predicted global patterns of scaling do not adequately explain the diversity in egg shapes. We show that egg size is not correlated with developmental rate and that, for many insects, egg size is not correlated with adult body size. Instead, we find that the evolution of parasitoidism and aquatic oviposition help to explain the diversification in the size and shape of insect eggs. Our study suggests that where eggs are laid, rather than universal allometric constants, underlies the evolution of insect egg size and shape. Size is a fundamental factor in many biological processes. The size of an 526 families and every currently described extant hexapod order24 organism may affect interactions both with other organisms and with (Fig. 1a and Supplementary Fig. 1). We combined this dataset with the environment1,2, it scales with features of morphology and physi- backbone hexapod phylogenies25,26 that we enriched to include taxa ology3, and larger animals often have higher fitness4. -

Proceedingscalif09cali
' BINDING LIST DEC 1 5 )9l1 Digitized by tine Internet Arciiive in 2009 with funding from Ontario Council of University Libraries http://www.archive.org/details/proceedingscalif09cali PROCEEDINGS California Academy of Sciences FOURTH SERIES Vol. IX 1919 o^n y ,% " PRINTED FROM THE ^• r JOHN W. HENDRIE PUBLICATION ENDOWMENT SAN FRANCISCO Published by the Academy 1919 r m COMMITTEE ON PUBLICATION George C. Edwards, Chairman C. E. Grunsky Barton Warren Evermann, Editor if CONTENTS OF VOLUME IX Plates 1-20 page Title-page ] Contents '" Notes on West American Chitons— II 1 By S. Stillman Berry (Published June 16, 1919) Life-Zone Indicators in California 37 By Harvey Monroe Hall and Joseph Grinnell (Published June 16, 1919) Notes on Mammals collected principally in Washington and California between the Years 1853 and 1874 by Dr. James Graham Cooper. ... 69 By Walter P. Taylor (Published July 12, 1919) Climatic Relations of the Tertiary and Quaternary Faunas of the California Region 123 By James Perrin Smith (Published July 12, 1919) Contribution to the Optics of the Microscope 175 By C. W. Woodworth (Published July 12, 1919) The Gopher-Snakes of Western North America 197 By John Van Denburgh and Joseph R. Slevin (Published August 26, 1919) New Oregon Diptera 221 By F. R. Cole and A. L. Lovett (Published August 26, 1919) Key to the North American Species of the Dipterous Genus Medeterus, with Descriptions of New Species 257 By Millard C. Van Duzee (Published August 26, 1919.1 Description of a New Fossil Fish from Japan 271 By David Starr Jordan (Published October 22, 1919) Notes on the Avifauna of the Inner Coast Range of California 273 By Joseph Mailliard (Published November 25, 1919) New Species of Flies (Diptera) from California 297 By J. -

Study of Systemic Status of Mycetophilidae
浙 江 林 学 院 学 报 2003 , 20(1):32 ~ 36 Journal of Zhejiang Forestry College Article ID :1000-5692(2003)01-0032-05 Study of systemic status of Mycetophilidae WANG Yi-ping , WU Hong , XU Hua-chao (Institute of Forest Protection , Zhejiang Forestry College, Linan 311300 , Zhejiang , China) Abstract :The actuality of Mycetophilidae family research has been illustrated in detail .The research includes biology , classification and idenfication , geographical distribution , status of systemic classification and system development .The existed problems and expectation are also explored in the paper .According to the latest information , Mycetophilidae family is composed of 5 subfamilies , i .e , Mycomyinae , Sciophilinae , Gnoristinae , Leiinae and Mycetophilinae .Mycetophilinae includes 2 tribes :Exechiini and Mycetophilini .References 28 Key words :Mycetophilidae ;geographical distribution ;classification status CLC Number :Q969.44 Document Code:A The family Mycetophilidae belongs to the superfamily Sciaroidea in the order Diptera , which is the largest family in the superfamily Sciaroidea[ 1] .Because of this insects feeding on edible fungi or large fungus body , a lot of scholars have been involved in the research on fungus pest species , and great progress in research on fungus gnats has been made .Based on research results and literature published at home and abroad in the past two decades , the authors make a summarization on the situation of the family Mycetophilidae insect species , including the biology , geography distribution , classification , phylogeny and make a forecast for the existing problems in the hope of providing some fundamental systemic materials for both theoretical research and practical application in the field of control of edible fungus pests[ 2 ~ 15] .