Alkaloids of Datura Innoxia Cardwellia Sublimis F
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Abelmoschus Moschatus Subsp
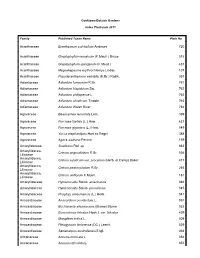
Cooktown Botanic Gardens Index Plantarum 2011 Family Published Taxon Name Plate No Acanthaceae Eranthemum pulchellum Andrews 720 Acanthaceae Graptophyllum excelsum (F.Meull.) Druce 515 Acanthaceae Graptophyllum spinigerum (F.Meull.) 437 Acanthaceae Megaskepasma erythrochlamys Lindau 107 Acanthaceae Pseuderanthemum variabile (R.Br.) Radlk. 357 Adiantaceae Adiantum formosum R.Br. 761 Adiantaceae Adiantum hispidulum Sw. 762 Adiantaceae Adiantum philippense L. 765 Adiantaceae Adiantum silvaticum Tindale 763 Adiantaceae Adiantum Walsh River 764 Agavaceae Beaucarnea recurvata Lem. 399 Agavaceae Furcraea foetida (L.) Haw. 637 Agavaceae Furcraea gigantea (L.) Haw. 049 Agavaceae Yucca elephantipes Hort.ex Regel 388 Agavaceae Agave sisalana Perrine. 159 Amarylidaceae Scadoxus Raf. sp 663 Amaryllidacea, Crinum angustifolium R.Br. 536 Liliaceae Amaryllidacea, Crinum asiaticum var. procerum (Herb. et Carey) Baker 417 Liliaceae Amaryllidacea, Crinum pedunculatum R.Br. 265 Liliaceae Amaryllidacea, Crinum uniflorum F.Muell. 161 Liliaceae Amaryllidaceae Hymenocallis Salisb. americanus 046 Amaryllidaceae Hymenocallis Salisb. peruvianna 045 Amaryllidaceae Proiphys amboinensis (L.) Herb. 041 Anacardiaceae Anacardium occidentale L. 051 Anacardiaceae Buchanania arborescens (Blume) Blume. 022 Anacardiaceae Euroschinus falcatus Hook.f. var. falcatus 429 Anacardiaceae Mangifera indica L. 009 Anacardiaceae Pleiogynium timorense (DC.) Leenh. 029 Anacardiaceae Semecarpus australiensis Engl. 368 Annonaceae Annona muricata L. 054 Annonaceae Annona reticulata L. 053 Annonaceae Annona squamosa 602 Annonaceae Cananga odorata (Lam.) Hook.f.&Thomson 406 Annonaceae Melodorum leichhardtii (F.Muell.) Diels. 360 Annonaceae Rollinia deliciosa Saff. 098 Apiaceae Centella asiatica (L.) Urb. 570 Apocynaceae Adenium obesum (Forssk.) Roem. & Schult. 489 Apocynaceae Allamanda cathartica L. 047 Apocynaceae Allamanda violacea Gardn. & Field. 048 Apocynaceae Alstonia actinophylla (A.Cunn.) K.Schum. 026 Apocynaceae Alstonia scholaris (L.) R.Br. 012 Apocynaceae Alyxia ruscifolia R.Br. -

Finschia-"A Genus of "Nut" Trees of the Southwest Pacific
Finschia-" A Genus of "Nut" Trees of the Southwest Pacific c. T. WHITE1 INTRODUCTION A PLANT FAMILY with a most interesting and F. Muell., Carnarvonia F. Muell., D arlin"gia F; intriguing distribution is Proteaceae, which finds Muell., Hollandaea F. Muell. (two spp.) , Mus its greatest development in Australia (650 " gravea F. Muell., and Placospermum White & species) on the one hand and South Africa (300 Francis. A surprising feature is the absence, species) on the other, though the two countries with the exception of one species in New Zea have no genera in common. Practically all the land, of the family "from Polynesia. South African species and the vast majority of There is in the islands of the southwest Paci "Australian ones are markedly xerophytic. The fic-Caroline Islands, New Guinea, Solomon largest genus, Greoillea R. Br., consists mainly Islands, and the New Hebrides-a group of trees of xerophytic shrubs or small trees but a few with the floral characters of Greuillea R. Br. are large trees found in the rain forests of and the fruit of Helicia Lour. These, I consider, tropical and subtropical eastern Australia, New all belong 'to Finschia Warb. This genus was Guinea, and New Caledonia. In the southwest founded by Warburg (1891: 297 ) on"a tree Pacific area the family finds its greatest develop from northeastern New Guinea. His original ment in northeastern Australia, where trees be description would cover Grevillea R. Br. exactly longing to it provide the great bulk of cabinet though he does not mention this genus and on timbers known in the trade as "Silky Oaks." the following page the distinctions he gives for There is close affinity between the Proteaceae of separating his proposed new genus from H elicia eastern Australia and of western South America are exactly those which distinguish Greuillea as illustrated by the genera Embothrium Forst. -

Grow Natives in Pots
ATG About the Garden ATGFACTAbout SHEET the Garden Fact Sheet No. 21 GROWING NATIVES IN POTS The key factors to successfully growing Australian plants in containers are exactly the same as for exotic plants - drainage and nutrition. The main points to remember are that good drainage is even more important for certain natives (mainly those originating in arid areas with very light, sandy soils) and that plants in the Proteaceae family require low phosphorus fertilisers. When growing plants in containers it is important *PLANTS IN THE to know their type and origin so ask yourself the following questions: FAMILY PROTEACEAE • Tree, shrub or climber/groundcover? • Does it originate in a high or low rainfall area? INCLUDE: • Is its natural habitat sun or shade? • Natural soil type - light (sandy) or heavy (clay)? Athertonia banksia The type of plant determines the ultimate pot size. buckinghamia Knowing its origins will tell you the type of growing cardwellia medium you need, how much water and whether it darlingia should be placed in sun or shade. If the label doesn’t dryandra give you enough information, ask nursery staff. hakea lomatia Any type of container can be used, providing it has macadamia plenty of adequate sized drainage holes. Too small and oreocallis they will drain too slowly and block up easily; too large persoonia and all your precious water will run away too fast. Most petrophile, good quality terracotta and plastic pots come with stenocarpus and good drainage holes; a medium-sized pot should have telopea no less than four, about the size of a five cent piece. -

Evolutionary History of Floral Key Innovations in Angiosperms Elisabeth Reyes
Evolutionary history of floral key innovations in angiosperms Elisabeth Reyes To cite this version: Elisabeth Reyes. Evolutionary history of floral key innovations in angiosperms. Botanics. Université Paris Saclay (COmUE), 2016. English. NNT : 2016SACLS489. tel-01443353 HAL Id: tel-01443353 https://tel.archives-ouvertes.fr/tel-01443353 Submitted on 23 Jan 2017 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. NNT : 2016SACLS489 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY, préparée à l’Université Paris-Sud ÉCOLE DOCTORALE N° 567 Sciences du Végétal : du Gène à l’Ecosystème Spécialité de Doctorat : Biologie Par Mme Elisabeth Reyes Evolutionary history of floral key innovations in angiosperms Thèse présentée et soutenue à Orsay, le 13 décembre 2016 : Composition du Jury : M. Ronse de Craene, Louis Directeur de recherche aux Jardins Rapporteur Botaniques Royaux d’Édimbourg M. Forest, Félix Directeur de recherche aux Jardins Rapporteur Botaniques Royaux de Kew Mme. Damerval, Catherine Directrice de recherche au Moulon Président du jury M. Lowry, Porter Curateur en chef aux Jardins Examinateur Botaniques du Missouri M. Haevermans, Thomas Maître de conférences au MNHN Examinateur Mme. Nadot, Sophie Professeur à l’Université Paris-Sud Directeur de thèse M. -

Fossil Fruit of the Macadamieae (Proteaceae) in the Tertiary of Eastern Australia: Eureka Gen
Memoirs of the Queensland Museum | Nature 55(1) © The State of Queensland (Queensland Museum) 2010 PO Box 3300, South Brisbane 4101, Australia Phone 06 7 3840 7555 Fax 06 7 3846 1226 Email [email protected] Website www.qm.qld.gov.au National Library of Australia card number ISSN 0079-8835 NOTE Papers published in this volume and in all previous volumes of the Memoirs of the Queensland Museum may be reproduced for scientific research, individual study or other educational purposes. Properly acknowledged quotations may be made but queries regarding the republication of any papers should be addressed to the Editor in Chief. Copies of the journal can be purchased from the Queensland Museum Shop. A Guide to Authors is displayed at the Queensland Museum web site http://www.qm.qld.gov.au/About+Us/Publications/Memoirs+of+the+Queensland+Museum A Queensland Government Project Typeset at the Queensland Museum Fossil fruit of the Macadamieae (Proteaceae) in the Tertiary of eastern Australia: Eureka gen. nov. Mary E. DETTMANN H. Trevor CLIFFORD Queensland Museum, Geosciences, Hendra Facility, 122 Gerler Rd, Hendra, Qld 4011, Australia. Email: [email protected] Citation: Dettmann, M.E. & Clifford, H.T. 2010 03 15. Fossil fruit of the Macadamieae (Proteaceae) in the Tertiary of eastern Australia: Eureka gen. nov.. Memoirs of the Queensland Museum — Nature 55(1): 147-166. Brisbane. ISSN 0079-8835. Accepted: 13 October 2009. ABSTRACT Eureka gen. nov. is proposed to accommodate fossil fruits recovered from several mid- Tertiary (early Oligocene-Miocene) sites in eastern Australia. The type (E. -

For Perspectives in Plant Ecology, Evolution and Systematics Manuscript Draft
Elsevier Editorial System(tm) for Perspectives in Plant Ecology, Evolution and Systematics Manuscript Draft Manuscript Number: PPEES-D-15-00109R1 Title: Bird pollinators, seed storage and cockatoo granivores explain large woody fruits as best seed defense in Hakea Article Type: Research paper Section/Category: Keywords: Black cockatoo; Crypsis; Fruit and seed size; Granivory; Resprouter; Spinescence Corresponding Author: Prof. Byron Lamont, Corresponding Author's Institution: Curtin University First Author: Byron Lamont Order of Authors: Byron Lamont; Byron Lamont; Mick Hanley; Philip Groom Abstract: Nutrient-impoverished soils with severe summer drought and frequent fire typify many Mediterranean-type regions of the world. Such conditions limit seed production and restrict opportunities for seedling recruitment making protection from granivores paramount. Our focus was on Hakea, a genus of shrubs widespread in southwestern Australia, whose nutritious seeds are targeted by strong-billed cockatoos. We assessed 56 Hakea species for cockatoo damage in 150 populations spread over 900 km in relation to traits expected to deter avian granivory: dense spiny foliage; large, woody fruits; fruit crypsis via leaf mimicry and shielding; low seed stores; and fruit clustering. We tested hypothesises centred on optimal seed defenses in relation to to a) pollination syndrome (bird vs insect), b) fire regeneration strategy (killed vs resprouting) and c) on-plant seed storage (transient vs prolonged). Twenty species in 50 populations showed substantial seed loss from cockatoo granivory. No subregional trends in granivore damage or protective traits were detected, though species in drier, hotter areas were spinier. Species lacking spiny foliage around the fruits (usually bird-pollinated) had much larger (4−5 times) fruits than those with spiny leaves and cryptic fruits (insect-pollinated). -

I Is the Sunda-Sahul Floristic Exchange Ongoing?
Is the Sunda-Sahul floristic exchange ongoing? A study of distributions, functional traits, climate and landscape genomics to investigate the invasion in Australian rainforests By Jia-Yee Samantha Yap Bachelor of Biotechnology Hons. A thesis submitted for the degree of Doctor of Philosophy at The University of Queensland in 2018 Queensland Alliance for Agriculture and Food Innovation i Abstract Australian rainforests are of mixed biogeographical histories, resulting from the collision between Sahul (Australia) and Sunda shelves that led to extensive immigration of rainforest lineages with Sunda ancestry to Australia. Although comprehensive fossil records and molecular phylogenies distinguish between the Sunda and Sahul floristic elements, species distributions, functional traits or landscape dynamics have not been used to distinguish between the two elements in the Australian rainforest flora. The overall aim of this study was to investigate both Sunda and Sahul components in the Australian rainforest flora by (1) exploring their continental-wide distributional patterns and observing how functional characteristics and environmental preferences determine these patterns, (2) investigating continental-wide genomic diversities and distances of multiple species and measuring local species accumulation rates across multiple sites to observe whether past biotic exchange left detectable and consistent patterns in the rainforest flora, (3) coupling genomic data and species distribution models of lineages of known Sunda and Sahul ancestry to examine landscape-level dynamics and habitat preferences to relate to the impact of historical processes. First, the continental distributions of rainforest woody representatives that could be ascribed to Sahul (795 species) and Sunda origins (604 species) and their dispersal and persistence characteristics and key functional characteristics (leaf size, fruit size, wood density and maximum height at maturity) of were compared. -

Ecology of Proteaceae with Special Reference to the Sydney Region
951 Ecology of Proteaceae with special reference to the Sydney region P.J. Myerscough, R.J. Whelan and R.A. Bradstock Myerscough, P.J.1, Whelan, R.J.2, and Bradstock, R.A.3 (1Institute of Wildlife Research, School of Biological Sciences (A08), University of Sydney, NSW 2006; 2Department of Biological Sciences, University of Wollongong, NSW 2522; 3Biodiversity Research and Management Division, NSW National Parks & Wildlife Service, PO Box 1967, Hurstville, NSW 1481) Ecology of Proteaceae with special reference to the Sydney region. Cunninghamia 6(4): 951–1015. In Australia, the Proteaceae are a diverse group of plants. They inhabit a wide range of environments, many of which are low in plant resources. They support a wide range of animals and other organisms, and show distinctive patterns of distribution in relation to soils, climate and geological history. These patterns of distribution, relationships with nutrients and other resources, interactions with animals and other organisms and dynamics of populations in Proteaceae are addressed in this review, particularly for the Sydney region. The Sydney region, with its wide range of environments, offers great opportunities for testing general questions in the ecology of the Proteaceae. For instance, its climate is not mediterranean, unlike the Cape region of South Africa, south- western and southern Australia, where much of the research on plants of Proteaceae growing in infertile habitats has been done. The diversity and abundance of Proteaceae vary in the Sydney region inversely with fertility of habitats. In the region’s rainforest there are few Proteaceae and their populations are sparse, whereas in heaths in the region, Proteaceae are often diverse and may dominate the canopy. -

Proteaceae Floral Crops; Cultivar Development and Underexploited Uses
Proteaceae Floral Crops; Cultivar Development and Underexploited Uses Kenneth W. Leonhardt and Richard A. Criley The Proteaceae apparently originated on the southern supercontinent Gondwana long before it divided and began drifting apart during the Mesozoic era, accounting for the presence of the Proteaceae on all of the southern continents (Brits 1984a). The Protea family comprises about 1400 species in over 60 genera, of which over 800 species in 45 genera are from Australia. Africa claims about 400 species, including 330 species in 14 genera from the western Cape. About 90 species occur in Central and South America, 80 on islands east of New Guinea, and 45 in New Caledonia. Madagascar, New Guinea, New Zealand, and South- east Asia host small numbers of species (Rebelo 1995). Proteas are neither herbaceous nor annual, and they are always woody. Their structural habit is vari- able from groundcover forms with creeping stems, and those with underground stems, to vertical to spread- ing shrubs, to tree forms. The leaves are generally large, lignified, hard, and leathery. A mature leaf will generally snap rather than fold when bent. The leaf anatomy is specially adapted for water conservation and drought resistance. These characteristics and the high leaf carbon to nitrogen ratio render the leaves indi- gestible to most insect pests (Rebelo 1995), accounting for the relatively pest-free status of most commer- cial protea plantings. The distribution of the family is linked to the occurrence of soils that are extremely deficient in plant nutrients (Brits 1984a). An accommodating characteristic of the family is the presence of proteoid roots. -

Contrasted Patterns of Hyperdiversification in Mediterranean Hotspots
Contrasted patterns of hyperdiversification in Mediterranean hotspots Herve´ Sauqueta,b,c,1, Peter H. Westonb, Cajsa Lisa Andersond, Nigel P. Barkere, David J. Cantrillc,f, Austin R. Mastg, and Vincent Savolainena,h aJodrell Laboratory, Royal Botanic Gardens, Kew, Richmond TW9 3DS, United Kingdom; bNational Herbarium of New South Wales, Botanic Gardens Trust, Mrs Macquaries Road, Sydney NSW 2000, Australia; cDepartment of Palaeobotany, Swedish Museum of Natural History, P.O. Box 50007, 104 05 Stockholm, Sweden; dDepartment of Systematic Biology, Evolutionary Biology Centre, Uppsala University, Norbyva¨gen 18D, SE-752 36 Uppsala, Sweden; eDepartment of Botany, Rhodes University, Grahamstown 6140, South Africa; fNational Herbarium of Victoria, Royal Botanic Gardens, Melbourne, Private Bag 2000, South Yarra, Victoria 3141, Australia; gDepartment of Biological Science, Florida State University, Tallahassee, FL 32306; and hImperial College London, Silwood Park Campus, Buckhurst Road, Ascot, Berkshire SL5 7PY, United Kingdom Edited by Peter R. Crane, University of Chicago, Chicago, IL, and approved November 12, 2008 (received for review June 9, 2008) Dating the Tree of Life has now become central to relating patterns Africa or Banksia in Western Australia. We use a rigorous of biodiversity to key processes in Earth history such as plate approach to select adequate calibration points and date a tectonics and climate change. Regions with a Mediterranean cli- multigene phylogenetic tree of all genera of this family. Prelim- mate have long been noted for their exceptional species richness inary phylogenetic analyses (12–15) have suggested that mem- and high endemism. How and when these biota assembled can bers of this group in each Mediterranean hotspot do not form a only be answered with a good understanding of the sequence of single clade but instead consist of multiple independent lineages, divergence times for each of their components. -

“Oak Tree”. the Genus Quercus Contains About 600 Species of Trees and Shrubs (The “True” Oaks) and Is Classified in the Family Fagaceae
Why are ‘Oaks’ called oaks? Quercus is Latin for “oak tree”. The Genus Quercus contains about 600 species of trees and shrubs (the “true” oaks) and is classified in the Family Fagaceae. This family includes Chestnuts (Castanea) the true Beeches (Fagus), the southern Beeches (Nothofagus) and other familiar trees. Quercus is native to the Northern Hemisphere, from cold latitudes to tropical Asia & the Americas. Leaves are spirally arranged, the fruit is an acorn and the genus contains both deciduous and evergreen (live) species. There are two main sub-genera: i) Quercus (with 4 or 5 Sections) and ii) Cyclobalanopsis which is not sub-divided. Within the Sub-genus Quercus, the four main groups are: a) Quercus – The white oaks of Europe, Asia and North America b) Cerris - Turkey oak & relatives in Europe & Asia (leaves with sharp lobes) c) Protobalanus – Canyon live oak & relatives in SW USA/Mexico (sharp lobes with bristles) d) Lobatae – Red (Black) oaks of North America & northern South America (sharp lobes with bristles) The Sub-genus Cyclobalanopsis contains evergreen, ring-cupped oaks of eastern & SE Asia. Botanical features of a tree can be used for identification (foliage, flower parts, fruit etc.), but they are of little value if all you have is a piece of wood. Thankfully, the anatomical features of the wood can also be used for identification and it is interesting to link the botanical features of a living tree to the anatomical features produced in the timber. The structure of hardwoods (Angiosperms) and softwoods (Conifers) differ significantly. Since Oaks are hardwoods, I will highlight the anatomical features of hardwoods. -

Cairns City in a Garden Master Plan 2007 #1987843 V1 © City of Cairns Table of Contents
CAIRNS CITY IN A GARDEN MASTER PLAN PART C PRECINCT PLANS 29 Cairns City in a Garden Master Plan 2007 #1987843 v1 © City of Cairns Table of Contents City in a Garden Master Plan Part C- Precinct Plans This section of the Cairns City in a Garden Master Plan provides a template for future tree planting in the key city streets (for a summary list of priority streets and tree species palettes refer to City in a Garden Master Plan Part E-Appendices) The precinct approach is in line with the Cairns Plan and addresses local issues and provides a guide to appropriate tree species selection for each precinct and outlines key objectives. For Central Cairns, the area defined by Florence Street to the north, the western footpath of the Esplanade to the east, the CBD side of the Wharf Street footpath to the south, and Bunda Street (east side of street) to the west, please refer to the Cairns CBD Master Plan 2007 for full planting details. CITY IN A GARDEN MASTER PLAN 2007 2 Vision Statement Error! Bookmark not defined. Context 2 CAIRNS CITY IN A GARDEN PART A-INTRODUCTION 10 CITY IN A GARDEN MASTER PLAN PART B-TREE SPECIES SELECTION 14 CITY IN A GARDEN MASTER PLAN PART C- PRECINCT PLANS 30 1. Cairns Beaches 32 1.1. Palm Cove 32 1.2. Clifton Beach 34 1.3. Kewarra Beach 36 1.4. Trinity Beach 38 2. Barron Smithfield 40 2.1. Trinity Park 40 2.2. Yorkeys Knob 42 2.3. Holloways Beach 44 Cairns City in a Garden 2.4.