Fossil Fruit of the Macadamieae (Proteaceae) in the Tertiary of Eastern Australia: Eureka Gen
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Macadamia Integrifolia HAES 741)
bioRxiv preprint doi: https://doi.org/10.1101/2020.05.25.114009; this version posted May 27, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Chromosome-scale assembly and annotation of the macadamia genome (Macadamia integrifolia HAES 741) Catherine J. Nock1†*, Abdul Baten1,2†, Ramil Mauleon1, Kirsty S. Langdon1, Bruce Topp3, Craig Hardner3, Agnelo Furtado3, Robert J. Henry3 and Graham J. King1 1Southern Cross Plant Science, Southern Cross University, Lismore NSW 2480, Australia 2AgResearch NZ, Grasslands Research Centre, Palmerston North 4442, New Zealand 3Queensland Alliance for Agriculture and Food Innovation, University of Queensland, St Lucia QLD 4069, Australia †These authors contributed equally to this work *Corresponding author, [email protected] ORCID 0000-0001-5609-4681 Abstract Macadamia integrifolia is a representative of the large basal eudicot family Proteaceae and the main progenitor species of the Australian native nut crop macadamia. Since its commercialisation in Hawaii fewer than 100 years ago, global production has expanded rapidly. However, genomic resources are limited in comparison to other horticultural crops. The first draft assembly of M. integrifolia had good coverage of the functional gene space but its high fragmentation has restricted its use in comparative genomics and association studies. Here we have generated an improved assembly of cultivar HAES 741 (4,094 scaffolds, 745 Mb, N50 413 kb) using a combination of Illumina paired and PacBio long read sequences. -

Method to Estimate Dry-Kiln Schedules and Species Groupings: Tropical and Temperate Hardwoods
United States Department of Agriculture Method to Estimate Forest Service Forest Dry-Kiln Schedules Products Laboratory Research and Species Groupings Paper FPL–RP–548 Tropical and Temperate Hardwoods William T. Simpson Abstract Contents Dry-kiln schedules have been developed for many wood Page species. However, one problem is that many, especially tropical species, have no recommended schedule. Another Introduction................................................................1 problem in drying tropical species is the lack of a way to Estimation of Kiln Schedules.........................................1 group them when it is impractical to fill a kiln with a single Background .............................................................1 species. This report investigates the possibility of estimating kiln schedules and grouping species for drying using basic Related Research...................................................1 specific gravity as the primary variable for prediction and grouping. In this study, kiln schedules were estimated by Current Kiln Schedules ..........................................1 establishing least squares relationships between schedule Method of Schedule Estimation...................................2 parameters and basic specific gravity. These relationships were then applied to estimate schedules for 3,237 species Estimation of Initial Conditions ..............................2 from Africa, Asia and Oceana, and Latin America. Nine drying groups were established, based on intervals of specific Estimation -

Brooklyn, Cloudland, Melsonby (Gaarraay)
BUSH BLITZ SPECIES DISCOVERY PROGRAM Brooklyn, Cloudland, Melsonby (Gaarraay) Nature Refuges Eubenangee Swamp, Hann Tableland, Melsonby (Gaarraay) National Parks Upper Bridge Creek Queensland 29 April–27 May · 26–27 July 2010 Australian Biological Resources Study What is Contents Bush Blitz? Bush Blitz is a four-year, What is Bush Blitz? 2 multi-million dollar Abbreviations 2 partnership between the Summary 3 Australian Government, Introduction 4 BHP Billiton and Earthwatch Reserves Overview 6 Australia to document plants Methods 11 and animals in selected properties across Australia’s Results 14 National Reserve System. Discussion 17 Appendix A: Species Lists 31 Fauna 32 This innovative partnership Vertebrates 32 harnesses the expertise of many Invertebrates 50 of Australia’s top scientists from Flora 62 museums, herbaria, universities, Appendix B: Threatened Species 107 and other institutions and Fauna 108 organisations across the country. Flora 111 Appendix C: Exotic and Pest Species 113 Fauna 114 Flora 115 Glossary 119 Abbreviations ANHAT Australian Natural Heritage Assessment Tool EPBC Act Environment Protection and Biodiversity Conservation Act 1999 (Commonwealth) NCA Nature Conservation Act 1992 (Queensland) NRS National Reserve System 2 Bush Blitz survey report Summary A Bush Blitz survey was conducted in the Cape Exotic vertebrate pests were not a focus York Peninsula, Einasleigh Uplands and Wet of this Bush Blitz, however the Cane Toad Tropics bioregions of Queensland during April, (Rhinella marina) was recorded in both Cloudland May and July 2010. Results include 1,186 species Nature Refuge and Hann Tableland National added to those known across the reserves. Of Park. Only one exotic invertebrate species was these, 36 are putative species new to science, recorded, the Spiked Awlsnail (Allopeas clavulinus) including 24 species of true bug, 9 species of in Cloudland Nature Refuge. -

IV. on the Proteaceć of Jussieu. by Mr. Robert Brown, Lib. LS
IV. On the Proteacea of Jussieu. By -Mr. Robert Brown, Lib. L.S. Read Jan. 17, 1809. THELinnean system of botany, though confessedly artificial, has not only contributed more than all others to facilitate tlie knowledge of species, but, by constantly directing the attention to those essential parts of the flower on which it is founded, has made us acquainted with more of their important modific-a t’ ions than we probably should have known, had it not been generally adopted, and has thus laid a more solid foundation for the esta- blishment of a natural arrangement, the superior importance of which no one has been inore fully impressed with than Linnzus hiinself. There are still, however, certain circumstances respccting the stamina and pistilla, which appear to iiie to havc been much less attended to than they deserve, both by Linneus and succeeding botanists. What I chiefly allude to is the state of these organs before the expansion of the flower. Tlie utility of ascertaining the internal condition of the ovarium before fecundation will liardly be called in question, now that the immortal worlis of Gxrtner and Jussieu hare demonstrated the necessity of minutely studying the fruits of plants in attempting to arrange tlicin ac- cording to tlic sum of their affinities, as in many cases the true nature of tlie ripc fruit, cspecially witli respect to the placenta- tion of the seeds, can oiily be detcrniined by this mc;~ns. Its importance is indeed expressly inculcated by many l~ot:inists, Tf’llO, 16 Mr. BROWN,on the Proteacee of Jussieu. -

Street Tree Master Plan Report © Sunshine Coast Regional Council 2009-Current
Sunshine Coast Street Tree Master Plan 2018 Part A: Street Tree Master Plan Report © Sunshine Coast Regional Council 2009-current. Sunshine Coast Council™ is a registered trademark of Sunshine Coast Regional Council. www.sunshinecoast.qld.gov.au [email protected] T 07 5475 7272 F 07 5475 7277 Locked Bag 72 Sunshine Coast Mail Centre Qld 4560 Acknowledgements Council wishes to thank all contributors and stakeholders involved in the development of this document. Disclaimer Information contained in this document is based on available information at the time of writing. All figures and diagrams are indicative only and should be referred to as such. While the Sunshine Coast Regional Council has exercised reasonable care in preparing this document it does not warrant or represent that it is accurate or complete. Council or its officers accept no responsibility for any loss occasioned to any person acting or refraining from acting in reliance upon any material contained in this document. Foreword Here on our healthy, smart, creative Sunshine Coast we are blessed with a wonderful environment. It is central to our way of life and a major reason why our 320,000 residents choose to live here – and why we are joined by millions of visitors each year. Although our region is experiencing significant population growth, we are dedicated to not only keeping but enhancing the outstanding characteristics that make this such a special place in the world. Our trees are the lungs of the Sunshine Coast and I am delighted that council has endorsed this master plan to increase the number of street trees across our region to balance our built environment. -
Abelmoschus Moschatus Subsp
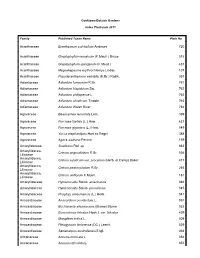
Cooktown Botanic Gardens Index Plantarum 2011 Family Published Taxon Name Plate No Acanthaceae Eranthemum pulchellum Andrews 720 Acanthaceae Graptophyllum excelsum (F.Meull.) Druce 515 Acanthaceae Graptophyllum spinigerum (F.Meull.) 437 Acanthaceae Megaskepasma erythrochlamys Lindau 107 Acanthaceae Pseuderanthemum variabile (R.Br.) Radlk. 357 Adiantaceae Adiantum formosum R.Br. 761 Adiantaceae Adiantum hispidulum Sw. 762 Adiantaceae Adiantum philippense L. 765 Adiantaceae Adiantum silvaticum Tindale 763 Adiantaceae Adiantum Walsh River 764 Agavaceae Beaucarnea recurvata Lem. 399 Agavaceae Furcraea foetida (L.) Haw. 637 Agavaceae Furcraea gigantea (L.) Haw. 049 Agavaceae Yucca elephantipes Hort.ex Regel 388 Agavaceae Agave sisalana Perrine. 159 Amarylidaceae Scadoxus Raf. sp 663 Amaryllidacea, Crinum angustifolium R.Br. 536 Liliaceae Amaryllidacea, Crinum asiaticum var. procerum (Herb. et Carey) Baker 417 Liliaceae Amaryllidacea, Crinum pedunculatum R.Br. 265 Liliaceae Amaryllidacea, Crinum uniflorum F.Muell. 161 Liliaceae Amaryllidaceae Hymenocallis Salisb. americanus 046 Amaryllidaceae Hymenocallis Salisb. peruvianna 045 Amaryllidaceae Proiphys amboinensis (L.) Herb. 041 Anacardiaceae Anacardium occidentale L. 051 Anacardiaceae Buchanania arborescens (Blume) Blume. 022 Anacardiaceae Euroschinus falcatus Hook.f. var. falcatus 429 Anacardiaceae Mangifera indica L. 009 Anacardiaceae Pleiogynium timorense (DC.) Leenh. 029 Anacardiaceae Semecarpus australiensis Engl. 368 Annonaceae Annona muricata L. 054 Annonaceae Annona reticulata L. 053 Annonaceae Annona squamosa 602 Annonaceae Cananga odorata (Lam.) Hook.f.&Thomson 406 Annonaceae Melodorum leichhardtii (F.Muell.) Diels. 360 Annonaceae Rollinia deliciosa Saff. 098 Apiaceae Centella asiatica (L.) Urb. 570 Apocynaceae Adenium obesum (Forssk.) Roem. & Schult. 489 Apocynaceae Allamanda cathartica L. 047 Apocynaceae Allamanda violacea Gardn. & Field. 048 Apocynaceae Alstonia actinophylla (A.Cunn.) K.Schum. 026 Apocynaceae Alstonia scholaris (L.) R.Br. 012 Apocynaceae Alyxia ruscifolia R.Br. -

Finschia-"A Genus of "Nut" Trees of the Southwest Pacific
Finschia-" A Genus of "Nut" Trees of the Southwest Pacific c. T. WHITE1 INTRODUCTION A PLANT FAMILY with a most interesting and F. Muell., Carnarvonia F. Muell., D arlin"gia F; intriguing distribution is Proteaceae, which finds Muell., Hollandaea F. Muell. (two spp.) , Mus its greatest development in Australia (650 " gravea F. Muell., and Placospermum White & species) on the one hand and South Africa (300 Francis. A surprising feature is the absence, species) on the other, though the two countries with the exception of one species in New Zea have no genera in common. Practically all the land, of the family "from Polynesia. South African species and the vast majority of There is in the islands of the southwest Paci "Australian ones are markedly xerophytic. The fic-Caroline Islands, New Guinea, Solomon largest genus, Greoillea R. Br., consists mainly Islands, and the New Hebrides-a group of trees of xerophytic shrubs or small trees but a few with the floral characters of Greuillea R. Br. are large trees found in the rain forests of and the fruit of Helicia Lour. These, I consider, tropical and subtropical eastern Australia, New all belong 'to Finschia Warb. This genus was Guinea, and New Caledonia. In the southwest founded by Warburg (1891: 297 ) on"a tree Pacific area the family finds its greatest develop from northeastern New Guinea. His original ment in northeastern Australia, where trees be description would cover Grevillea R. Br. exactly longing to it provide the great bulk of cabinet though he does not mention this genus and on timbers known in the trade as "Silky Oaks." the following page the distinctions he gives for There is close affinity between the Proteaceae of separating his proposed new genus from H elicia eastern Australia and of western South America are exactly those which distinguish Greuillea as illustrated by the genera Embothrium Forst. -

Pathogens Associated with Diseases. of Protea, Leucospermum and Leucadendron Spp
PATHOGENS ASSOCIATED WITH DISEASES. OF PROTEA, LEUCOSPERMUM AND LEUCADENDRON SPP. Lizeth Swart Thesis presented in partial fulfillment of the requirements for the degree of Master of Science in Agriculture at the University of Stellenbosch Supervisor: Prof. P. W. Crous Decem ber 1999 Stellenbosch University https://scholar.sun.ac.za DECLARATION 1, the undersigned, hereby declare that the work contained in this thesis is my own original work and has not previously in its entirety or in part been submitted at any university for a degree. SIGNATURE: DATE: Stellenbosch University https://scholar.sun.ac.za PATHOGENS ASSOCIATED WITH DISEASES OF PROTEA, LEUCOSPERMUM ANDLEUCADENDRONSPP. SUMMARY The manuscript consists of six chapters that represent research on different diseases and records of new diseases of the Proteaceae world-wide. The fungal descriptions presented in this thesis are not effectively published, and will thus be formally published elsewhere in scientific journals. Chapter one is a review that gives a detailed description of the major fungal pathogens of the genera Protea, Leucospermum and Leucadendron, as reported up to 1996. The pathogens are grouped according to the diseases they cause on roots, leaves, stems and flowers, as well as the canker causing fungi. In chapter two, several new fungi occurring on leaves of Pro tea, Leucospermum, Telopea and Brabejum collected from South Africa, Australia or New Zealand are described. The following fungi are described: Cladophialophora proteae, Coniolhyrium nitidae, Coniothyrium proteae, Coniolhyrium leucospermi,Harknessia leucospermi, Septoria prolearum and Mycosphaerella telopeae spp. nov. Furthermore, two Phylloslicla spp., telopeae and owaniana are also redecribed. The taxonomy of the Eisinoe spp. -

Approved NSW & National Recovery Plan Eidothea Hardeniana
Approved NSW & National Recovery Plan Eidothea hardeniana September 2004 © Department of Environment and Conservation (NSW), July 2004. This work is copyright. However, material presented in this plan may be copied for personal use or published for educational purposes, providing that any extracts are fully acknowledged. Apart from this and any other use as permitted under the Copyright Act 1968, no part may be reproduced without prior written permission from NSW Department of Environment and Conservation. NSW Department of Environment and Conservation 43 Bridge Street (PO Box 1967) Hurstville NSW 2220 Tel: 02 9585 6444 www.nationalparks.nsw.gov.au Requests for information or comments regarding the recovery program for the Nightcap Oak are best directed to: The Nightcap Oak Recovery Co-ordinator Threatened Species Unit, North East Branch NSW Department of Environment and Conservation Locked Bag 914 Coffs Harbour NSW 2450 Tel: 02 6651 5946 Cover illustrator: Lesley Elkan © Botanic Gardens Trust, Sydney Cover illustration: Adult and juvenile leaves and fruit of Eidothea hardeniana This plan should be cited as follows: NSW Department of Environment and Conservation 2004, Recovery Plan for the Nightcap Oak (Eidothea hardeniana), Department of Environment and Conservation (NSW), Hurstville. ISBN 0 7313 6781 2 Recovery Plan The Nightcap Oak Recovery Plan for the Nightcap Oak (Eidothea hardeniana) Foreword The New South Wales Government established a new environment agency on 24 September 2003, the Department of Environment and Conservation (NSW), which incorporates the New South Wales National Parks and Wildlife Service. Responsibility for the preparation of Recovery Plans now rests with this new department. This document constitutes the New South Wales State and National Recovery Plan for Eidothea hardeniana Weston & Kooyman (Nightcap Oak), and as such considers the conservation requirements of the species across its range. -

Plant Life of Western Australia
INTRODUCTION The characteristic features of the vegetation of Australia I. General Physiography At present the animals and plants of Australia are isolated from the rest of the world, except by way of the Torres Straits to New Guinea and southeast Asia. Even here adverse climatic conditions restrict or make it impossible for migration. Over a long period this isolation has meant that even what was common to the floras of the southern Asiatic Archipelago and Australia has become restricted to small areas. This resulted in an ever increasing divergence. As a consequence, Australia is a true island continent, with its own peculiar flora and fauna. As in southern Africa, Australia is largely an extensive plateau, although at a lower elevation. As in Africa too, the plateau increases gradually in height towards the east, culminating in a high ridge from which the land then drops steeply to a narrow coastal plain crossed by short rivers. On the west coast the plateau is only 00-00 m in height but there is usually an abrupt descent to the narrow coastal region. The plateau drops towards the center, and the major rivers flow into this depression. Fed from the high eastern margin of the plateau, these rivers run through low rainfall areas to the sea. While the tropical northern region is characterized by a wet summer and dry win- ter, the actual amount of rain is determined by additional factors. On the mountainous east coast the rainfall is high, while it diminishes with surprising rapidity towards the interior. Thus in New South Wales, the yearly rainfall at the edge of the plateau and the adjacent coast often reaches over 100 cm. -

Rates of Molecular Evolution and Diversification in Plants: Chloroplast
Duchene and Bromham BMC Evolutionary Biology 2013, 13:65 http://www.biomedcentral.com/1471-2148/13/65 RESEARCH ARTICLE Open Access Rates of molecular evolution and diversification in plants: chloroplast substitution rates correlate with species-richness in the Proteaceae David Duchene* and Lindell Bromham Abstract Background: Many factors have been identified as correlates of the rate of molecular evolution, such as body size and generation length. Analysis of many molecular phylogenies has also revealed correlations between substitution rates and clade size, suggesting a link between rates of molecular evolution and the process of diversification. However, it is not known whether this relationship applies to all lineages and all sequences. Here, in order to investigate how widespread this phenomenon is, we investigate patterns of substitution in chloroplast genomes of the diverse angiosperm family Proteaceae. We used DNA sequences from six chloroplast genes (6278bp alignment with 62 taxa) to test for a correlation between diversification and the rate of substitutions. Results: Using phylogenetically-independent sister pairs, we show that species-rich lineages of Proteaceae tend to have significantly higher chloroplast substitution rates, for both synonymous and non-synonymous substitutions. Conclusions: We show that the rate of molecular evolution in chloroplast genomes is correlated with net diversification rates in this large plant family. We discuss the possible causes of this relationship, including molecular evolution driving diversification, speciation increasing the rate of substitutions, or a third factor causing an indirect link between molecular and diversification rates. The link between the synonymous substitution rate and clade size is consistent with a role for the mutation rate of chloroplasts driving the speed of reproductive isolation. -

Grow Natives in Pots
ATG About the Garden ATGFACTAbout SHEET the Garden Fact Sheet No. 21 GROWING NATIVES IN POTS The key factors to successfully growing Australian plants in containers are exactly the same as for exotic plants - drainage and nutrition. The main points to remember are that good drainage is even more important for certain natives (mainly those originating in arid areas with very light, sandy soils) and that plants in the Proteaceae family require low phosphorus fertilisers. When growing plants in containers it is important *PLANTS IN THE to know their type and origin so ask yourself the following questions: FAMILY PROTEACEAE • Tree, shrub or climber/groundcover? • Does it originate in a high or low rainfall area? INCLUDE: • Is its natural habitat sun or shade? • Natural soil type - light (sandy) or heavy (clay)? Athertonia banksia The type of plant determines the ultimate pot size. buckinghamia Knowing its origins will tell you the type of growing cardwellia medium you need, how much water and whether it darlingia should be placed in sun or shade. If the label doesn’t dryandra give you enough information, ask nursery staff. hakea lomatia Any type of container can be used, providing it has macadamia plenty of adequate sized drainage holes. Too small and oreocallis they will drain too slowly and block up easily; too large persoonia and all your precious water will run away too fast. Most petrophile, good quality terracotta and plastic pots come with stenocarpus and good drainage holes; a medium-sized pot should have telopea no less than four, about the size of a five cent piece.