Pine Defoliator Bupalus Piniaria L. (Lepidoptera: Geometridae) and Its Entomopathogenic Fungi 1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

E-News Winter 2019/2020
Winter e-newsletter December 2019 Photos Merry Christmas and a Happy New Year! INSIDE THIS ISSUE: Contributions to our newsletters Dates for your Diary & Winter Workparties....2 Borage - Painted Lady foodplant…11-12 are always welcome. Scottish Entomological Gathering 2020 .......3-4 Lunar Yellow Underwing…………….13 Please use the contact details Obituary - David Barbour…………..………….5 Chequered Skipper Survey 2020…..14 below to get in touch! The Bog Squad…………………………………6 If you do not wish to receive our Helping Hands for Butterflies………………….7 newsletter in the future, simply Munching Caterpillars in Scotland………..…..8 reply to this message with the Books for Sale………………………...………..9 word ’unsubscribe’ in the title - thank you. RIC Project Officer - Job Vacancy……………9 Coul Links Update……………………………..10 VC Moth Recorder required for Caithness….10 Contact Details: Butterfly Conservation Scotland t: 01786 447753 Balallan House e: [email protected] Allan Park w: www.butterfly-conservation.org/scotland Stirling FK8 2QG Dates for your Diary Scottish Recorders’ Gathering - Saturday, 14th March 2020 For everyone interested in recording butterflies and moths, our Scottish Recorders’ Gathering will be held at the Battleby Conference Centre, by Perth on Saturday, 14th March 2020. It is an opportunity to meet up with others, hear all the latest butterfly and moth news and gear up for the season to come! All welcome - more details will follow in the New Year! Highland Branch AGM - Saturday, 18th April 2020 Our Highlands & Island Branch will be holding their AGM on Saturday, 18th April in a new venue, Green Drive Hall, 36 Green Drive, Inverness, IV2 4EU. More details will follow on the website in due course. -

Conservation Assessment for the Kansan Spikerush Leafhopper (Dorydiella Kansana Beamer)
Conservation Assessment For The Kansan spikerush leafhopper (Dorydiella kansana Beamer) USDA Forest Service, Eastern Region January 11, 2005 James Bess OTIS Enterprises 13501 south 750 west Wanatah, Indiana 46390 This document is undergoing peer review, comments welcome This Conservation Assessment was prepared to compile the published and unpublished information on the subject taxon or community; or this document was prepared by another organization and provides information to serve as a Conservation Assessment for the Eastern Region of the Forest Service. It does not represent a management decision by the U.S. Forest Service. Though the best scientific information available was used and subject experts were consulted in preparation of this document, it is expected that new information will arise. In the spirit of continuous learning and adaptive management, if you have information that will assist in conserving the subject taxon, please contact the Eastern Region of the Forest Service - Threatened and Endangered Species Program at 310 Wisconsin Avenue, Suite 580 Milwaukee, Wisconsin 53203. TABLE OF CONTENTS EXECUTIVE SUMMARY ............................................................................................................ 1 ACKNOWLEDGEMENTS............................................................................................................ 1 NOMENCLATURE AND TAXONOMY ..................................................................................... 1 DESCRIPTION OF SPECIES....................................................................................................... -
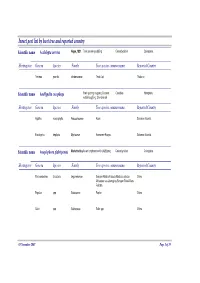
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

64 Part I. Description and Backgrounds of a Polymorphism in the Pine Looper
64 PART I. DESCRIPTION AND BACKGROUNDS OF A POLYMORPHISM IN THE PINE LOOPER 1. THE OCCURRENCE OF A YELLOW-GREEN POLYMORPHISM IN PINE LOOPER CATERPILLARS IN THE NETHERLANDS 1.1. SOME GENERAL REMARKS ON THE OBJECTS OF STUDY The pine looper, or bordered white, is the only species of the genus Bupalus belonging to the lepidopterous family of the Geometridae. In Europe this moth has been an object of study since the last century, due to its being a very serious pest insect in some parts of the vast area in which it is found. Its food consists of needles of Scots pine, Pinus sylvestris L. The adult has a wing span of 3.5-4.0 cm. The male is dark brown with white or cream patches on the wings. The female has a greyish brown or an orange brown ground colour. Her markings are more or less similar to those of the male, though less distinct. The eggs are green, about 1 mm long, oval, with a shallow depression at the upper side. The first two larval instars are yellowish green, the first instar having a brown head, and the second instar a yellowish brown head. From the third instar onwards the larvae are green, with several disruptive white or yellowish longitudinal stripes both on the body and the head. An extensive description of the larvae has been given by HERREBOUT, KUYTEN & DE RUITER (1963). The pupae are green during the first few days following pupation, after which they become dark brown. The species is univoltine. The moths emerge from the end of May to the end of June. -

Newsletter 90
Norfolk Moth Survey c/o Natural History Dept., Castle Museum, Norwich, NR1 3JU Newsletter No.90 November 2016 INTRODUCTION With the flurry of activity through the latter part of the summer, it is easy to forget how cool, wet and frustrating the early part of the season often was. Opinion generally seems to suggest that, while the range of species seen was much to be expected, actual numbers of moths were down on the whole. However, one event during that early period brought the subject of moths to the attention of the media, both locally and nationally. This was the great invasion of Diamond- backed moths, Plutella xylostella, that took place at the very end of May and the first days of June. It would be no exaggeration to say that literally millions of these tiny moths arrived on these shores, with at least one commentator describing it as “...a plague of biblical proportion”. Several of us found ourselves answering queries and calls from a variety of sources in connection with this influx. Despite the dire warnings proffered by some sections of the media - and others, our cabbages weren’t totally obliterated as a result. In fact, the expected boost in numbers resulting from these original invaders breeding here, just didn’t seem to happen. In what might have otherwise been a distinctly average season, it is good to be able to report that twelve new species have been added to the Norfolk list this year. Amazingly, seven of these have been adventives, including one species new for the UK. -

The Moths of Dunnet Forest Caithness
THE MOTHS OF CAITHNE SS THE MOTHS OF DUNNET F O R E S T CAITHNESS A REVIEW OF THE MOTH SPECIES RECORDED IN DUNNET FOREST CAITHNESS AS AT THE ST 31 DECEMBER 2018 N E I L M O N E Y COUNTY MOTH RECORDER CAITHNESS PURPOSE OF THIS REVIEW The development of Dunnet Forest from the original Forestry Commission experimental coniferous woodland to a community woodland managed by the community through the Dunnet Forestry Trust has reached a stage where changes in habitat are starting to drive change in the moth species being recorded in the forest. The purpose of this review is to detail moth species recorded up to 31st December 2018 to provide a base line against which future change can be monitored BACKGROUND The original forest was planted by the Forestry Commission in the mid-1950s as an experiment in forestry planting on poor soils. The forest was acquired by Scottish Natural Heritage in 1984 and is part of the Dunnet Links Site of Special Scientific Interest. Since 2003 the forest has been under the management of the Dunnet Forestry Trust (DFT) a community trust run by volunteers and employing two part time professional foresters. Covering 104 hectares the original coniferous planting was of a range of species but dominated by Sitka Spruce, Lodgepole Pine, Corsican Pine and Mountain Pine. About half of the area developed into mature forest with the remainder become a mixture of open space, scattered trees and scrub woodland.1 A CHANGING HABITAT DFT have been pro-active in the managing the forest both as a recreational resource for the community and in diversifying the habitat. -

Tarset and Greystead Biological Records
Tarset and Greystead Biological Records published by the Tarset Archive Group 2015 Foreword Tarset Archive Group is delighted to be able to present this consolidation of biological records held, for easy reference by anyone interested in our part of Northumberland. It is a parallel publication to the Archaeological and Historical Sites Atlas we first published in 2006, and the more recent Gazeteer which both augments the Atlas and catalogues each site in greater detail. Both sets of data are also being mapped onto GIS. We would like to thank everyone who has helped with and supported this project - in particular Neville Geddes, Planning and Environment manager, North England Forestry Commission, for his invaluable advice and generous guidance with the GIS mapping, as well as for giving us information about the archaeological sites in the forested areas for our Atlas revisions; Northumberland National Park and Tarset 2050 CIC for their all-important funding support, and of course Bill Burlton, who after years of sharing his expertise on our wildflower and tree projects and validating our work, agreed to take this commission and pull everything together, obtaining the use of ERIC’s data from which to select the records relevant to Tarset and Greystead. Even as we write we are aware that new records are being collected and sites confirmed, and that it is in the nature of these publications that they are out of date by the time you read them. But there is also value in taking snapshots of what is known at a particular point in time, without which we have no way of measuring change or recognising the hugely rich biodiversity of where we are fortunate enough to live. -

Moths and Butterflies
LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004MOTHS LJL©2004 LJL©2004AND BUTTERFLIES LJL©2004 LJL©2004 (LEPIDOPTERA) LJL©2004 LJL©2004 LJL©2004 FROM LJL©2004 BAHÍA LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004HONDA LJL©2004 LJL©2004 AND CANALES LJL©2004 LJL©2004 DE TIERRA LJL©2004 ISLANDLJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004(VERAGUAS, LJL©2004 LJL©2004 PANAMA LJL©2004) LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 LJL©2004 -

Current Problems and Tasks of Forest Protection in Poland
Folia Forestalia Polonica, Series A – Forestry, 2018, Vol. 60 (3), 161–172 ORIGINAL ARTICLE DOI: 10.2478/ffp-2018-0016 Current problems and tasks of forest protection in Poland Iwona Skrzecz1 , Aldona Perlińska2 1 Forest Research Institute, Department of Forest Protection, Sękocin Stary, Braci Leśnej 3, 05-090 Raszyn, Poland, phone: +48 22 7150541, e-mail: [email protected] 2 General Directorate of the State Forests, Department of Forest Protection, Grójecka 127, 02-124 Warszawa, Poland AbstrAct Current problems of forest protection concern the declining health of forest stands due to climate change and the re- sulting extreme weather events such as heat waves, droughts, hurricane winds, heavy rainfalls and floods. Repeated impacts of these factors increase susceptibility of forest stands to pest insects and fungal pathogens. Norway spruce (Picea abies [L.] H. Karst) is sensitive to high air temperatures and water shortage. Long lasting droughts during the last two decades, have been one of the reasons behind Norway spruce dieback due to severe outbreak of European spruce bark beetle Ips typographus (L.) in the mountainous regions of southern Poland. In the Scots pine (Pinus sylvestris L.) stands, water balance disorders have enhanced the colonization of weekend trees by steelblue jewel beetle Phaenops cyanea (F.) and engraver beetle Ips acuminatus (Gyll.), as well as contributed to the spread of fungal diseases caused by Gremmeniella abietina (Lagerb.) Cenangium ferruginosum Fr. and Sphaeropsis sapinea Fr. fungi. Water related stress leads to weakening of oak stands, which are attacked by Agrilus spp. beetles and pathogens from the genus Phytophthora. It is possible that long lasting droughts initiated the spread of infectious ash disease caused by Hymenoscyphus fraxineus (= Chalara fraxinea), which resulted in the epidemic of ash dieback throughout Europe. -

Mid-Atlantic Forest Ecosystem Vulnerability Assessment and Synthesis: a Report from the Mid-Atlantic Climate Change Response Framework Project
United States Department of Agriculture Mid-Atlantic Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Mid-Atlantic Climate Change Response Framework Project Forest Northern General Technical Service Research Station Report NRS-181 October 2018 ABSTRACT Forest ecosystems will be affected directly and indirectly by a changing climate over the 21st century. This assessment evaluates the vulnerability of 11 forest ecosystems in the Mid-Atlantic region (Pennsylvania, New Jersey, Delaware, eastern Maryland, and southern New York) under a range of future climates. We synthesized and summarized information on the contemporary landscape, provided information on past climate trends, and described a range of projected future climates. This information was used to parameterize and run multiple forest impact models, which provided a range of potential tree responses to climate. Finally, we brought these results before two multidisciplinary panels of scientists and land managers familiar with the forests of this region to assess ecosystem vulnerability through a formal consensus-based expert elicitation process. Each chapter of this assessment builds on the previous chapter. The description of the contemporary landscape presents major forest trends and stressors currently threatening forests in the Mid-Atlantic region and defines the forest communities being assessed. The background information in Chapter 2 summarizes climate data analysis and climate models. Analysis of climate records in Chapter 3 indicates that average temperatures and total precipitation in the region have increased. Downscaled climate models in Chapter 4 project potential increases in temperature in every season, but projections for precipitation indicate slight increases in winter and spring, and high variability in summer and fall projections, depending on the scenario. -

Molecular Phylogenetics and Evolution 162 (2021) 107198
Molecular Phylogenetics and Evolution 162 (2021) 107198 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Molecular phylogeny, classification, biogeography and diversification patterns of a diverse group of moths (Geometridae: Boarmiini) a,b,* c d ~ e,f g Leidys Murillo-Ramos , Nicolas Chazot , Pasi Sihvonen , Erki Ounap , Nan Jiang , Hongxiang Han g, John T. Clarke e,h, Robert B. Davis e, Toomas Tammaru e, Niklas Wahlberg a a Department of Biology, Lund University, Lund, Sweden b Departamento de Biología, Universidad de Sucre, Sucre, Colombia c Department of Ecology, Swedish University of Agricultural Sciences, Uppsala, Sweden d Finnish Museum of Natural History, Helsinki, Finland e Department of Zoology, Institute of Ecology and Earth Sciences, University of Tartu, Tartu, Estonia f Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Tartu, Estonia g Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing, China h Department of Ecology and Biogeography, Faculty of Biological and Veterinary Sciences, Nicolaus Copernicus University, Lwowska, Torun,´ Poland ARTICLE INFO ABSTRACT Keywords: Understanding how and why some groups have become more species-rich than others, and how past biogeog Lepidoptera raphy may have shaped their current distribution, are questions that evolutionary biologists have long attempted polyphagyPolyphagy to answer. We investigated diversification patterns and historical biogeography of a hyperdiverse lineage of female flightlessness Lepidoptera, the geometrid moths, by studying its most species-rich tribe Boarmiini, which comprises ca. 200 boarmiines genera and ca. known 3000 species. We inferred the evolutionary relationships of Boarmiini based on a dataset of Cleora Biston 346 taxa, with up to eight genetic markers under a maximum likelihood approach. -

Newsletter 16 - 2011
Butterfly Conservation Highland Branch - Newsletter 16 - 2011 Butterfly Conservation Saving butterflies, moths and our environment Butterfly Conservation Highland Branch Newsletter 16 Spring 2011 www.highland-butterflies.org.uk 1 Butterfly Conservation Highland Branch - Newsletter 16 - 2011 2 Butterfly Conservation Highland Branch - Newsletter 16 - 2011 Contents Introduction 4 Disused roads, mini nature reserves. 5 Small Blues at Loch Fleet. 6 Memorable moments, 2010. 8 Polmaily scrub bash. 9 Ruby Tiger bites back. 11 Peacocks in Ardnamurchan. 12 Hungry Caterpillars turn out to be the Brightest Butterflies 13 Branch membership - latest numbers and distribution 13 Report from South Uist. 14 2010 Moth Summary - Shetland. 15 Moths Count in Wester Ross. 16 Report from the West Coast 2010. 18 Macro-moth records from East Inverness-shire. 22 Some moth highlights from Moray. 23 Lepidoptera in Banffshire 2010. 25 Moths of Caithness website. 35 Rearing Moths. 36 Scottish Macro-moth List 2011. 38 Lifetime Achievement in Lepidoptera Conservation. 38 Moth publications. 39 Butterfly publications. 40 Obituary: Barbara Ballinger; Derek C Hume. 41 Wider Countryside Butterfly Survey. 42 Which moth species are pollinating Lesser Butterfly Orchid in Scotland? 43 Your butterfly records 44 Comma survey to be launched. 44 Branch Committee, Butterfly Recorders, County Moth Recorders. 45 Highland Branch Field Trips 2011. 47 3 Butterfly Conservation Highland Branch - Newsletter 16 - 2011 that involvement in voluntary biological record- ing.” What struck a chord with me as I ran through the questions and responses were how very many of the various elements I was interested in. A mish-mash of subjects and interests, the sum total of which gives one a tremendous return for one’s efforts.