Canadian Manuscript Report of Fisheries and Aquatic Scienes No
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sailing Directions (Enroute)
PUB. 154 SAILING DIRECTIONS (ENROUTE) ★ BRITISH COLUMBIA ★ Prepared and published by the NATIONAL GEOSPATIAL-INTELLIGENCE AGENCY Bethesda, Maryland © COPYRIGHT 2007 BY THE UNITED STATES GOVERNMENT NO COPYRIGHT CLAIMED UNDER TITLE 17 U.S.C. 2007 TENTH EDITION For sale by the Superintendent of Documents, U.S. Government Printing Office Internet: http://bookstore.gpo.gov Phone: toll free (866) 512-1800; DC area (202) 512-1800 Fax: (202) 512-2250 Mail Stop: SSOP, Washington, DC 20402-0001 Preface 0.0 Pub. 154, Sailing Directions (Enroute) British Columbia, 0.0NGA Maritime Domain Website Tenth Edition, 2007, is issued for use in conjunction with Pub. http://www.nga.mil/portal/site/maritime 120, Sailing Directions (Planning Guide) Pacific Ocean and 0.0 Southeast Asia. Companion volumes are Pubs. 153, 155, 157, 0.0 Courses.—Courses are true, and are expressed in the same 158, and 159. manner as bearings. The directives “steer” and “make good” a 0.0 Digital Nautical Chart 26 provides electronic chart coverage course mean, without exception, to proceed from a point of for the area covered by this publication. origin along a track having the identical meridianal angle as the 0.0 This publication has been corrected to 21 July 2007, includ- designated course. Vessels following the directives must allow ing Notice to Mariners No. 29 of 2007. for every influence tending to cause deviation from such track, and navigate so that the designated course is continuously Explanatory Remarks being made good. 0.0 Currents.—Current directions are the true directions toward 0.0 Sailing Directions are published by the National Geospatial- which currents set. -

Pacific Region 1994 Management Plan Geoduck and Horse Clam
PACIFIC REGION 1994 MANAGEMENT PLAN GEODUCK AND HORSE CLAM 1'hiJ Geoduclc and Horse QIJm MtJMgement Plan is intended for general puposes only. When there is a discrepancy between the PlDn and the regulations. the regulations are the final aurhority. description of Areas and Subareas referenced in this Plan can be found in the Pacific Fishery Management Area !guJations. Fisheries Peches and Oceans et Oceans Canada 1. TABLE OF CONTENTS 2. DFO Contacts for Further Information/Geoduck Observers . .. 3 3. SheUfish - Goals and Objectives ..................... ,...................... 4 4. Enforcement Plan .............•...............•......•................. 5 5. Aboriginal Fisheries Strstegy .......•....•.•.•.•.•.•....................... 5 6. Sport Fishing . • . .. 6 7. History of the Horse Clam Fishery . • . • . • . • . • • . .. 6 8. Horse Clam Commercial Fishing Plan - 1994 ...............•.................... 7 8.1. Management Changes ..........................•................. 7 8.2. Seasons, Quotas and Open Times •.. • . • . .. 7 8.3. Permanent Ares Closures . • . • . • . .. 8 8.4. Ucencing.................................................... 8 8.4.1. Ucence Category ........................................ 8 8.4.2. Species .............................................. 8 8.4.3. Gear ............................•................... 8 8.5. Catch and Fisbing Data ........................................... 8 9. History of the Geoduck Fishery . .. 8 10. Geoduck Commercial Fishing Plan - 1994 . • . .. 9 10.1. Management Changes . ... .. 9 -

2015 Annual Report
Albert Myshrall, a wood worker for many years, taught himself how to make hand-crafted banjos and ukeleles. The instruments he crafts from his home in Queen Charlotte City exhibit superior workmanship and sound quality and are showcased in Northern Development’s shop local program. www.lovehaidagwaii.com | Photo credit: Jason Shafto CONTENTS ABOUT THE TRUST .............................. 4 PERFORMANCE INDICATORS .............. 51 Service Region ................................................................ 6 FUND MANAGEMENT ......................... 55 By the Numbers .............................................................. 7 Mission, Values, Ethics Statements .................................. 8 Northwest Readiness ....................................................... 57 Message from the chair and CEO .................................... 10 Connecting bc ................................................................ 58 NCLGA ........................................................................... 58 GOVERNANCE ...................................... 12 Efficient Fund Management and Catalyst 2 ..................... 59 Board of Directors ........................................................... 13 BEST PRACTICES .................................. 60 Regional Advisory Committees ........................................ 14 2015 FUND BALANCES ........................ 67 FINANCIAL OVERVIEW ........................ 16 INDEPENDENT AUDITORS REPORT ..... 72 STRATEGIC PLAN AT A GLANCE .......... 19 PROGRAM DELIVERY ......................... -

Energy Resources in BC's Central Interior
Western Geography, 12 (2002), pp. 216–229 ©Western Division, Canadian Association of Geographers Energy Resources in BC’s Central Interior Peter Ostergaard Chairperson and Chief Executive Officer British Columbia Utilities Commission Reliable, convenient, and reasonably priced energy services are a prerequisite for both quality of life and an energy-intensive resource sector in central British Columbia. Investments by governments, Crown corpo- rations, and BC-based companies in energy infra- structure to facilitate regional economic development characterized most of the last 50 years. This is chang- ing. While access to infrastructure remains important, the region’s gas and electricity price advantages are dis- appearing as these commodities are traded across conti- nental infrastructure networks. Future energy invest- ments in the region will be more environmentally benign and perhaps environmentally beneficial, as efforts to continue to mitigate the environmental impacts of reservoirs, reservoir operations, and energy corridors. Introduction The Central Interior’s abundance of low cost energy resources has strongly influenced the evolution of the region’s economy and patterns of settlement. Aside from biomass supplies (e.g., firewood, pulping liquor, hog fuel), the region imports most of its energy needs using a highly developed and reliable network of oil and gas pipelines and power transmission lines. Access to these supplies has been a prerequisite to attracting the energy intensive, resource based industries upon which the region will continue to depend. However, these abundant, low cost supplies have come at an envi- ronmental cost, primarily in the form of flooded valley bottoms and land alienated by pipeline and transmission corridors. Energy Resources in BC’s Central Interior 217 Historical Context Traditionally when we think of energy, its sources come to mind. -

Regular Council Meeting
AGENDA for the Council Meeting to be held on Monday, April 24, 2017 in the Council Chambers of the Municipal Office, 1 Mackenzie Boulevard, Mackenzie, BC. CALLED TO ORDER 7:15 PM 1. ADOPTION OF MINUTES a) Committee Meeting - April 10, 2017. 7 - 8 b) Regular Meeting - April 10, 2017. 9 - 12 2. INTRODUCTION OF LATE ITEMS 3. ADOPTION OF AGENDA 4. PETITIONS AND DELEGATIONS a) Regional District of Fraser-Fort George Jim Martin, Chief Administrative Officer, and Michael Higgins, Emergency Services Coordinator, will be presenting to Council in regards to their Region Wide Emergency Preparedness Initiative. b) Regional District of Fraser-Fort George Petra Wildauer, General Manager of Environmental Services, will present to Council in regards to the construction of a full service transfer station at the Mackenzie Regional Landfill. 5. CORRESPONDENCE Motion required to accept all correspondence listed on the agenda. April 24, 2017 For Action: a) Letter from the Mackenzie Outdoor Route and Trail 13 Association (MORATA) requesting that Council provide them with the following resolution of support for their grant application to the Northern Development Initiative Trust for their Morfee Mountain Bike Trail Development Phase One Project: "THAT the District of Mackenzie Council supports the application to the Northern Development Initiative Trust from the Mackenzie Outdoor Route Trail Association for a grant up to $30,000 for the Morfee Mountain Bike Trail Development Phase One project from the Prince George Development Account." b) Letter from the Mackenzie -

Conservation and Sustainable Forestry Plan TFL 54 June 2005
Conservation and Sustainable Forestry Plan TFL 54 June 2005 CONSERVATION AND SUSTAINABLE FORESTRY PLAN DRAFT MANAGEMENT PLAN No.4 TREE FARM LICENSE 54 Effective January 1 2005 to December 31, 2009. Submitted ____________________, 2005. International Forest Products Ltd. Authors Gordon Butt, Madrone Environmental Services J. Dan Lousier, Whiskeyjack Science Don McMillan, Interfor Warren Warttig, Interfor DRAFT Interfor Madrone Conservation and Sustainable Forestry Plan TFL 54 June 2005 EXECUTIVE SUMMARY Like ordinary TFL Management Plans, this Conservation and Sustainable Forestry Plan will show how Interfor will manage TFL 54 in Clayoquot Sound over the coming five-year period (2005 to 2009). However, Clayoquot Sound is not an ordinary place and this is not an ordinary Management Plan. This Plan describes the special context and history of the Sound, it explains how Interfor has met the challenges of the last decade, and will show how Interfor will continue to sustainably manage the TFL in the future. The 1995 Clayoquot Sound Scientific Panel recommendations resulted in fundamental and dramatic changes and marked a clear departure from earlier, conflict-inspiring industrial forest management focusing on timber production. The preparation of this plan marks the 10th anniversary since Interfor began the implementation of the Scientific Panel’s recommendations. In fully embracing these changes, Interfor has been on the forefront of ecosystem-based management in BC. It now has 10 years experience working with the new way of doing business. Interfor has adjusted to a new philosophy that emphasizes the trees left behind, rather than the trees harvested. Interfor can claim numerous successes. It has: 1. -
Public Accounts of British Columbia Fiscal Year Ended March 31
9345 00412867 STACKS HJ 13 B561 1969/70 c. 1 SIMON FRASER UNIVERSITY LIBRARY FOR USE IN LIBRARY ONLY Digitized by the Internet Archive in 2019 with funding from University of Alberta Libraries https://archive.org/details/publicaccountsof00mini_0 i I *7 '/ P'-'i* PROVINCE OF BRITISH COLUMBIA PUBLIC ACCOUNTS FISCAL YEAR ENDED MARCH 31, 1970 PERIOD FROM APRIL 1, 1969, TO MARCH 31, 1970 Printed by A. Sutton, Printer to the Queen’s Most Excellent Majesty in right of the Province of British Columbia. 1970 To Colonel the Honourable John R. Nicholson, P.C., O.B.E., Q.C., LL.D., Lieutenant-Governor of the Province of British Columbia. May it please Your Honour: The undersigned has the honour to present the Public Accounts of the Province of British Columbia for the fiscal year ended March 31, 1970. W. A. C. BENNETT, Minister of Finance. Department of Finance, Victoria, British Columbia, May 15, 1970. Victoria, British Columbia, May 15, 1970. The Honourable W. A. C. Bennett, P.C., LL.D., D.Pol.Sc., K.St.J., Minister of Finance. Sir,—I have the honour to submit herewith the Public Accounts of the Prov¬ ince of British Columbia for the fiscal year ended March 31, 1970. Respectfully submitted. C. J. FERBER, C.A., Comptroller-General. PUBLIC ACCOUNTS, 1969/70 E 5 Order of Public Accounts Main Statements: Ref. No. Statement of Financial Condition as at March 31, 1970_ E 6-7 Revenue and Expenditure (Current Account, Summary by Departments) for the Fiscal Year Ended March 31, 1970_ E 8-9 Cash Receipts and Payments (Consolidated Revenue Fund) for the Fiscal -

The Scrivener Fall 2018
Fall 2018 | Volume 27 Number 3 Published Quarterly by The Society of Notaries Public of British Columbia 27 years Morrie Bailey Daniel Boisvert Lorne Mann Jacqui Mendes Introducing the Board of the BC Notaries Association inside: Town and Country BC Notaries and Allied Professionals Publications Mail Agreement: 40010827 At Work around Our Province ST_partnerAd_Scrivener_8.38x10.75_v2_final.pdf 1 9/27/18 4:38 PM Creative Adaptable Analytical Today’s successful real estate professional is committed to lifelong learning. UBC’s Continuing Professional Development Courses meet your needs. • Convenient online course format • Broad range of valuation and real estate business topics • Guaranteed Appraisal Institute of Canada’s Continuing Professional Development credits Business Development Series: Eight courses designed to provide practitioners with the entrepreneurial and practical skills to establish and successfully operate a real estate business. Entrepreneurship and Small Business Development Business Strategy: Managing a Profitable Real Estate Business Succession Planning for Real Estate Professionals Organizing and Financing a Real Estate Business Accounting and Taxation Considerations for a Real Estate Business Marketing and Technology Considerations for a Real Estate Business Human Resources Management Considerations in Real Estate Law and Ethical Considerations in Real Estate Business To find out more, visit: realestate.ubc.ca/CPD tel: 604.822.2227 / 1.877.775.7733 email: [email protected] Scrivener-2018.indd 1 2/22/2018 -

Schedule of Indian Reserves and Settlements
Schedule of Indian Reserves and Settlements Schedule of Indian Reserves and Settlements Preface 1. This schedule contains lists of Indian Reserves and Settlements in Canada by Provinces and lists of Indian Bands by Regions taken from information filed with the Indian Land Registry as of June 1, 1972. 2. Indian Settlements are shown in the "Reserve" columns of Parts 1 and 2. These are not Indian Reserves within the meaning of the Indian Act and occupation of settlements is not restricted to a particular Band. 3. Part 1 of the schedule lists Indian Reserves and Settlements alphabetically by Provinces. Settlements follow after the alphabetical list of Reserves for each Province. This section also states the names of the Indian Bands for whose use and benefit the Reserves have been set apart, the approximate geographical location and the approximate acreage of each Reserve. 4. Map designations are given in the location description. The N.T.S. abbreviation means the "National Topographical Systems" of maps covering the Reserves and adjacent land. Maps can be obtained by writing to the Map Distribution Office, Department of Energy, Mines and Resources, Ottawa, Ontario. When ordering maps the complete N.T.S. designations must be quoted. To assist in locating Reserve location, approximate degrees and minutes latitude and longitude have been given. 5. Acreage includes all lands in which the Band has a recorded interest. Acreages are shown as APPROXIMATE because in most cases they are based on old data which have not yet been verified. Research and surveys to establish accurate acreage are being carried out and the acreage shown may be changed in the future. -
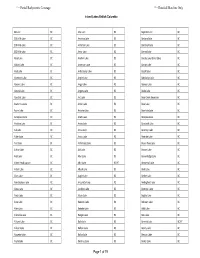
^ = Partial Bathymetric Coverage * = Detailed Shoreline Only Page 1 of 19
^ = Partial Bathymetric Coverage * = Detailed Shoreline Only Inland Lakes British Columbia #3 Lake BC Alta Lake BC Baptiste Lake BC 103 Mile Lake BC Amanita Lake BC Barbara Lake BC 108 Mile Lake BC Ambrose Lake BC Bardolph Lake BC 130 Mile Lake BC Amor Lake BC Barnes Lake BC Abas Lake BC Anahim Lake BC Barsby Lake (Blind Lake) BC Abbott Lake BC Anderson Lake BC Barton Lake BC Abel Lake BC Andy Bailey Lake BC Basalt Lake BC Aberdeen Lake BC Angler Lake BC Battleship Lake BC Abrams Lake BC Angly Lake BC Baynes Lake BC Abruzzi Lake BC Angora Lake BC Beale Lake BC Abuntlet Lake BC Ant Lake BC Bear Creek Reservoir BC Academus Lake BC Antler Lake BC Bear Lake BC Acorn Lake BC Antoine Lake BC Bearhole Lake BC Aeroplane Lake BC Anutz Lake BC Bearpaw Lake BC Ahdatay Lake BC Anzac Lake BC Beartrack Lake BC Aid Lake BC Anzus Lake BC Beartrap Lake BC Aiken Lake BC Arctic Lake BC Beatrice Lake BC Aird Lake BC Armstrong Lake BC Beaux Yeux Lake BC Airline Lake BC Ash Lake BC Beaver Lake BC Alah Lake BC Atan Lake BC Beaverlodge Lake BC Albert Head Lagoon BC Atlin Lake BC/YT Beavertail Lake BC Albert Lake BC Atluck Lake BC Beck Lake BC Alces Lake BC Augier Lake BC Becker Lake BC Alex Graham Lake BC Azouzetta Lake BC Bedingfield Lake BC Alexis Lake BC Azuklotz Lake BC Bednesti Lake BC Aleza Lake BC Azure Lake BC Begbie Lake BC Alice Lake BC Babcock Lake BC Belcourt Lake BC Allan Lake BC Babette Lake BC Bells Lake BC Allendale Lake BC Badger Lake BC Ben Lake BC Alleyne Lake BC Baile Lake BC Bennett Lake BC/YT Allison Lake BC Balfour Lake BC Benny Lake BC -

Lumber & Veneer, for BC
THE BRITISH COLUMBIA RAILWAY AND REGIONAL DEVELOPMENT by ELLSWORTH PAUL GAMBLE B.A., University of Washington, 1969 A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF ARTS in the School of Community and Regional Planning We accept this thesis as conforming to the required standard THE UNIVERSITY OF BRITISH COLUMBIA April, 1972 In presenting this thesis in partial fulfilment of the requirements for an advanced degree at the University of British Columbia, I agree that the Library shall make it freely available for reference and study. I further agree that permission for extensive copying of this thesis for scholarly purposes may be granted by the Head of my Department or by his representatives. It is understood that copying or publication of this thesis for financial gain shall not be allowed without my written permission. Department of Community and Regional Planning The University of British Columbia Vancouver 8, Canada Date April 28f 1972 ii. ' ABSTRACT This thesis considers the Pacific Great Eastern Railway, the British Columbia Railway as of April 1, 1972, a Provincial Crown corpor• ation, and the implications of its extensions upon regional development. The indicators of regional development studied are population and indust• rial profiles. The time framework of the thesis is from 1952 to 1972, the period of the P.G.E. extensions. Two perspectives of the implications for regional development are examined. Chapter Two treats with the Provincial Government agencies whose policies have had the most effect in the study area. The financing, safety, and freight rate implications of the P.G.E. -

2017 Williston Fish Hg Report June 2018
Williston-Dinosaur Watershed Fish Mercury Investigation 2017 Report Prepared for: Fish and Wildlife Compensation Program, Peace Region 3333 22nd Ave. Prince George, BC V2N 1B4 June 2018 Azimuth Consulting Group Partnership 218-2902 West Broadway Vancouver BC, V6K 2G8 Project No. CO94394 Williston-Dinosaur Watershed Fish Mercury Investigation – 2017 Report EXECUTIVE SUMMARY The Fish and Wildlife Compensation Program (FWCP) – Peace Region carried out a strategic planning process in 2012-13 to review and identify program priorities in this region. Guided by a Strategic Planning Group (SPG), including First Nations, academia, BC Hydro and the FWCP-Peace Board, a Peace Basin Plan and six Action Plans were finalized in 2014. Objective 3a of the Reservoirs Action Plan is to “Improve understanding of mercury concentrations, contamination pathways and potential effects on human health and the broader ecosystem.” Initial efforts on this objective were commissioned by FWCP Peace in 2014 and identified the need to obtain updated information on fish mercury concentrations and consumption habits. In 2016, the Azimuth Consulting Group (Azimuth) team (including EDI Environmental Dynamics [EDI], Chu Cho Environmental [CCE] and Hagen and Associates) was awarded a multi-year contract to collect fish mercury data from the Parsnip, Peace, Finlay reaches of Williston and Dinosaur reservoirs and reference lakes (i.e., the Williston-Dinosaur Watershed Fish Mercury Study). Results of this investigation will assess provide an updated fish mercury database for the Williston-Dinosaur watershed and understanding of how results compare with nearby reference lakes. The long-term goal is to ‘update’ the existing fish consumption advisory, in partnership with provincial health agencies.