Environmental Health Criteria 94 Permethrin
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cypermethrin
International Environmental Health Criteria 82 Cypermethrin Published under the joint sponsorship of the United Nations Environment Programme, the International Labour Organisation, and the World Health Organization WORLD HEALTH ORGANIZATION GENEVA 1989 Other titles available in the ENVIRONMENTAL HEALTH CRITERIA series include: 1. Mercury 2. Polychlorinated Biphenyls and Terphenyls 3. Lead 4. Oxides of Nitrogen 5. Nitrates, Nitrites, and N-Nitroso Compounds 6. Principles and Methods for Evaluating the Toxicity of Chemicals, Part 1 7. Photochemical Oxidants 8. Sulfur Oxides and Suspended Particulate Matter 9. DDT and its Derivatives 10. Carbon Disulfide 11. Mycotoxins 12. Noise 13. Carbon Monoxide 14. Ultraviolet Radiation 15. Tin and Organotin Compounds 16. Radiofrequency and Microwaves 17. Manganese 18. Arsenic 19. Hydrogen Sulfide 20. Selected Petroleum Products 21. Chlorine and Hydrogen Chloride 22. Ultrasound 23. Lasers and Optical Radiation 24. Titanium 25. Selected Radionuclides 26. Styrene 27. Guidelines on Studies in Environmental Epidemiology 28. Acrylonitrile 29. 2,4-Dichlorophenoxyacetic Acid (2,4-D) 30. Principles for Evaluating Health Risks to Progeny Associated with Exposure to Chemicals during Pregnancy 31. Tetrachloroethylene 32. Methylene Chloride 33. Epichlorohydrin 34. Chlordane 35. Extremely Low Frequency (ELF) Fields 36. Fluorine and Fluorides 37. Aquatic (Marine and Freshwater) Biotoxins 38. Heptachlor 39. Paraquat and Diquat 40. Endosulfan 41. Quintozene 42. Tecnazene 43. Chlordecone 44. Mirex continued on p. 156 -

1 Chiral Analysis of Pollutants and Their Metabolites by Capillary
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Digital.CSIC Chiral analysis of pollutants and their metabolites by capillary electromigration methods Javier Hernández-Borges1, Miguel Ángel Rodríguez-Delgado1, Francisco J. García-Montelongo1, Alejandro Cifuentes2,* 1Department of Analytical Chemistry, Nutrition and Food Science, University of La Laguna, Avda. Astrofísico Fco. Sánchez s/n, 38071 La Laguna, Tenerife, Spain. 2 Department of Food Analysis, Institute of Industrial Fermentations (CSIC), Juan de la Cierva 3, 28006 Madrid, Spain. Keywords: capillary electrophoresis; enantiomers; pollutants; pesticides; review; chiral analysis; CE; MEKC; CEC. Corresponding author: Dr. Alejandro Cifuentes E-mail: [email protected] Fax: +34-91-5644853; Tel: +34-91-5622900 1 Abbreviations: allyl-TER: 1-allylterguride; ANDSA: 7-aminonaphthalene-1,3-disulfonic acid; ANSA: 5-aminonaphthalene-1-sulfonic acid; ANTS: 8-aminonaphthalene-1,3,6-trisulfonic acid; BGE: background electrolyte; CM-γ-CD: carboxymethylated-γ-cyclodextrin; MCPA: (4-chloro-2-methylphenoxy)-acetic acid; MCPB: (4-chloro-2-methylphenoxy)butyric acid; 2,2-CPPA: 2-(2-chlorophenoxy)-propionic acid; 2,3-CPPA: 2-(3- chlorophenoxy)propionic acid; 2,4-CPPA: 2-(4-chlorophenoxy)-propionic acid; CMBA: 2-(4-chlorophenyl)-3-methylbutanoic acid; CA: chrysanthemic acid; MEGA: decanoyl-N-methylglucamide; DCA: dichorochrysanthemic acid; 2,4-D: (2,4- dichlorophenoxy)acetic acid; 2,4-DB: 4-(2,4-dichlorophenoxy)butiric acid; 2,4- DCPPA: 2-(2,4-dichlorophenoxy)propionic -

Genetically Modified Baculoviruses for Pest
INSECT CONTROL BIOLOGICAL AND SYNTHETIC AGENTS This page intentionally left blank INSECT CONTROL BIOLOGICAL AND SYNTHETIC AGENTS EDITED BY LAWRENCE I. GILBERT SARJEET S. GILL Amsterdam • Boston • Heidelberg • London • New York • Oxford Paris • San Diego • San Francisco • Singapore • Sydney • Tokyo Academic Press is an imprint of Elsevier Academic Press, 32 Jamestown Road, London, NW1 7BU, UK 30 Corporate Drive, Suite 400, Burlington, MA 01803, USA 525 B Street, Suite 1800, San Diego, CA 92101-4495, USA ª 2010 Elsevier B.V. All rights reserved The chapters first appeared in Comprehensive Molecular Insect Science, edited by Lawrence I. Gilbert, Kostas Iatrou, and Sarjeet S. Gill (Elsevier, B.V. 2005). All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopy, recording, or any information storage and retrieval system, without permission in writing from the publishers. Permissions may be sought directly from Elsevier’s Rights Department in Oxford, UK: phone (þ44) 1865 843830, fax (þ44) 1865 853333, e-mail [email protected]. Requests may also be completed on-line via the homepage (http://www.elsevier.com/locate/permissions). Library of Congress Cataloging-in-Publication Data Insect control : biological and synthetic agents / editors-in-chief: Lawrence I. Gilbert, Sarjeet S. Gill. – 1st ed. p. cm. Includes bibliographical references and index. ISBN 978-0-12-381449-4 (alk. paper) 1. Insect pests–Control. 2. Insecticides. I. Gilbert, Lawrence I. (Lawrence Irwin), 1929- II. Gill, Sarjeet S. SB931.I42 2010 632’.7–dc22 2010010547 A catalogue record for this book is available from the British Library ISBN 978-0-12-381449-4 Cover Images: (Top Left) Important pest insect targeted by neonicotinoid insecticides: Sweet-potato whitefly, Bemisia tabaci; (Top Right) Control (bottom) and tebufenozide intoxicated by ingestion (top) larvae of the white tussock moth, from Chapter 4; (Bottom) Mode of action of Cry1A toxins, from Addendum A7. -

TITLE: Lindane and Other Treatments for Lice and Scabies: a Review of Clinical Effectiveness and Safety
TITLE: Lindane and Other Treatments for Lice and Scabies: A Review of Clinical Effectiveness and Safety DATE: 11 June 2010 CONTEXT AND POLICY ISSUES: Head lice infestation (Pediculosis capitis) affects millions of children and adults worldwide each year.1 Direct head-to-head contact is the most common mode of transmission.2 The highest prevalence of infestation occurs in school aged children aged three to eleven years, with girls being more commonly affected than boys.1,2 Although head lice are not generally associated with serious morbidity, they are responsible for significant social embarrassment and lost productivity in schools or offices.1 Scabies, an infestation of the skin by the mite Sarcoptes scabiei, represents a common public health concern particularly in overcrowded communities with a high prevalence of poverty.3 Scabies is transmitted by close-person contact and occasionally by clothing or linens.3 Complications include secondary bacterial infections and post-streptococcal glomerulonephritis.3 Topical products available in Canada for the treatment of head lice and scabies are presented in Appendix 1 and Appendix 2. Insecticidal agents such as permethrin and lindane have historically been considered the standard treatments for head lice and scabies.2,3 Toxicity is low following topical administration of permethrin due to minimal percutaneous absorption.4 However, several jurisdictions have banned lindane due to concerns of neurotoxicity and bone marrow suppression, as well as potential negative effects on the environment (contamination of waste water).5 Furthermore, widespread use of permethrin, pyrethrins/piperonyl butoxide, and lindane has led to resistance and higher rates of treatment failure.6 Resistance patterns and rates to these agents in Canada have not yet been studied.6 Due to concerns surrounding resistance and neurotoxicity, patients and caregivers have searched for alternative treatments. -
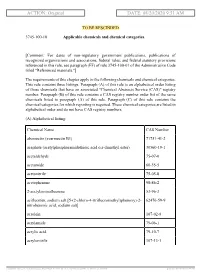
ACTION: Original DATE: 08/20/2020 9:51 AM
ACTION: Original DATE: 08/20/2020 9:51 AM TO BE RESCINDED 3745-100-10 Applicable chemicals and chemical categories. [Comment: For dates of non-regulatory government publications, publications of recognized organizations and associations, federal rules, and federal statutory provisions referenced in this rule, see paragraph (FF) of rule 3745-100-01 of the Administrative Code titled "Referenced materials."] The requirements of this chapter apply to the following chemicals and chemical categories. This rule contains three listings. Paragraph (A) of this rule is an alphabetical order listing of those chemicals that have an associated "Chemical Abstracts Service (CAS)" registry number. Paragraph (B) of this rule contains a CAS registry number order list of the same chemicals listed in paragraph (A) of this rule. Paragraph (C) of this rule contains the chemical categories for which reporting is required. These chemical categories are listed in alphabetical order and do not have CAS registry numbers. (A) Alphabetical listing: Chemical Name CAS Number abamectin (avermectin B1) 71751-41-2 acephate (acetylphosphoramidothioic acid o,s-dimethyl ester) 30560-19-1 acetaldehyde 75-07-0 acetamide 60-35-5 acetonitrile 75-05-8 acetophenone 98-86-2 2-acetylaminofluorene 53-96-3 acifluorfen, sodium salt [5-(2-chloro-4-(trifluoromethyl)phenoxy)-2- 62476-59-9 nitrobenzoic acid, sodium salt] acrolein 107-02-8 acrylamide 79-06-1 acrylic acid 79-10-7 acrylonitrile 107-13-1 [ stylesheet: rule.xsl 2.14, authoring tool: RAS XMetaL R2_0F1, (dv: 0, p: 185720, pa: -

(12) United States Patent (10) Patent No.: US 8,852,618 B2 Clough (45) Date of Patent: Oct
USOO8852618B2 (12) United States Patent (10) Patent No.: US 8,852,618 B2 Clough (45) Date of Patent: Oct. 7, 2014 (54) INSECTICIDAL MIXTURE CONTAINING CA 2429218 A1 6, 2002 GAMMA-CYHALOTHRN CH 689326 A5 4f1995 EP O237227 A1 9, 1987 EP 0771526 A2 5, 1997 (75) Inventor: Martin Stephen Clough, Bracknell EP O988788 A1 3f2000 (GB) FR 272O230 A1 12/1995 JP 63. 126805 A2 5, 1988 (73) Assignee: Syngenta Limited, Guildford (GB) JP 63126805 A2 5, 1988 JP 63126805 5, 1998 c - r WO WO 86 O7525 A1 12, 1986 (*) Notice: Subject to any disclaimer, the term of this WO WO 93 03618 A2 3, 1993 patent is extended or adjusted under 35 WO WO95 229O2 A1 8/1995 U.S.C. 154(b) by 824 days. WO WO9533380 A1 12, 1995 WO WO 96 16543 A2 6, 1996 (21) Appl. No.: 12/633,063 WO WO97 06687 A1 2/1997 WO WO974O692 A1 11, 1997 (22) Filed: Dec.a V88, 2009 WO WOOOO2453 A1 1, 2000 OTHER PUBLICATIONS (65) Prior Publication Data US 201O/OO81714 A1 Apr. 1, 2010 Canadian Office Action (Applin. No. 2,452,515 filed: Jul. 10, 2002) mailing date Oct. 1, 2010 (pp. 1-2). Related U.S. Application Data Allen et al. Transgenic & Conventional Insect & Weed Control Sys tems; Proceedings of the Beltwide Cotton Conference, vol. 2, 1065 (62) Division of application No. 10/484.745, filed as 1068 (1999), USA. application No. PCT/GB02/03181 on Jul. 10, 2002, Anonymous; Pesticide Mixtures for Control of Insect and Acarid now Pat. No. -

NASTTPO CFATS Information 2Nd Edition
National Association of SARA Title III Program Officials Concerned with the Emergency Planning and Community Right-to-Know Act January 13, 2008 Issue 07-01 2nd Edition The Department of Homeland Security has adopted 6 CFR Part 27, a new regulation mandated by Congress, known as Chemical Facility Anti-Terrorism Standards (CFATS). The regulation is intended to fill a security gap in our country’s anti-terrorism efforts by identifying and improving the security of chemicals that are potentially at a high level of risk for release, theft, or sabotage. LEPCs and SERCs should alert EPCRA & RMP reporting facilities about these requirements. No reports are due to the LEPCs and SERCs under these requirements; however, given the potential for security requirements to have an impact on facility access for emergency responders and on emergency plans, it is critical for local planners, responders and facilities to communicate in order for a facility to satisfy the regulatory requirements. In order to aid LEPCs, SERCs and facilities in understanding these new requirements we have prepared some short-hand aids. Following this cover page is a key issue comparison between EPCRA, RMP and the CFATS regulation. As requirements may change the user is counseled to look for updated information. Following the side-by-side comparison we have edited the EPA “List of Lists” to add the Appendix A list of chemicals and thresholds from the CFATS program. This list may change and we will update these materials when that happens. The initial requirement for a facility with an Appendix A chemical at or over the threshold is to submit a Top-Screen. -

Table II. EPCRA Section 313 Chemical List for Reporting Year 2017 (Including Toxic Chemical Categories)
Table II. EPCRA Section 313 Chemical List For Reporting Year 2017 (including Toxic Chemical Categories) Individually listed EPCRA Section 313 chemicals with CAS numbers are arranged alphabetically starting on page II-3. Following the alphabetical list, the EPCRA Section 313 chemicals are arranged in CAS number order. Covered chemical categories follow. Note: Chemicals may be added to or deleted from the list. The Emergency Planning and Community Right-to-Know Call Center or the TRI-Listed Chemicals website will provide up-to-date information on the status of these changes. See section B.3.c of the instructions for more information on the de minimis % limits listed below. There are no de minimis levels for PBT chemicals since the de minimis exemption is not available for these chemicals (an asterisk appears where a de minimis limit would otherwise appear in Table II). However, for purposes of the supplier notification requirement only, such limits are provided in Appendix C. Chemical Qualifiers Certain EPCRA Section 313 chemicals listed in Table II have parenthetic “qualifiers.” These qualifiers indicate that these EPCRA Section 313 chemicals are subject to the section 313 reporting requirements if manufactured, processed, or otherwise used in a specific form or when a certain activity is performed. An EPCRA Section 313 chemical that is listed without a qualifier is subject to reporting in all forms in which it is manufactured, processed, and otherwise used. The following chemicals are reportable only if they are manufactured, processed, or otherwise used in the specific form(s) listed below: Chemical/ Chemical Category CAS Number Qualifier Aluminum (fume or dust) 7429-90-5 Only if it is a fume or dust form. -
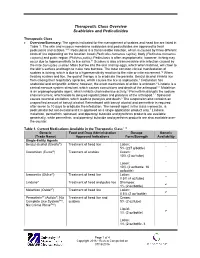
Therapeutic Class Overview Scabicides and Pediculicides
Therapeutic Class Overview Scabicides and Pediculicides Therapeutic Class Overview/Summary: The agents indicated for the management of scabies and head lice are listed in Table 1. The skin and mucous membrane scabicides and pediculicides are approved to treat pediculosis and scabies.1-10 Pediculosis is a transmissible infection, which is caused by three different kinds of lice depending on the location: head (Pediculus humanus capitis), body (Pediculus humanus corporis) and pubic region (Phthirus pubis). Pediculosis is often asymptomatic; however, itching may occur due to hypersensitivity to lice saliva.11 Scabies is also a transmissible skin infection caused by the mite Sarcoptes scabiei. Mites burrow into the skin and lay eggs, which when hatched, will crawl to the skin’s surface and begin to make new burrows. The most common clinical manifestation of scabies is itching, which is due to a hypersensitivity reaction to the mite or mite excrement.12 When treating scabies and lice, the goal of therapy is to eradicate the parasite. Benzyl alcohol inhibits lice from closing their respiratory spiracles, which causes the lice to asphyxiate.3 Crotamiton has scabicidal and antipruritic actions; however, the exact mechanism of action is unknown.4 Lindane is a central nervous system stimulant, which causes convulsions and death of the arthropod.1,2 Malathion is an organophosphate agent, which inhibits cholinesterase activity.5 Permethrin disrupts the sodium channel current, which leads to delayed repolarization and paralysis of the arthropod.1,2 Spinosad causes neuronal excitation, which leads to paralysis and death.6 The suspension also contains an unspecified amount of benzyl alcohol. -

Estonian Statistics on Medicines 2016 1/41
Estonian Statistics on Medicines 2016 ATC code ATC group / Active substance (rout of admin.) Quantity sold Unit DDD Unit DDD/1000/ day A ALIMENTARY TRACT AND METABOLISM 167,8985 A01 STOMATOLOGICAL PREPARATIONS 0,0738 A01A STOMATOLOGICAL PREPARATIONS 0,0738 A01AB Antiinfectives and antiseptics for local oral treatment 0,0738 A01AB09 Miconazole (O) 7088 g 0,2 g 0,0738 A01AB12 Hexetidine (O) 1951200 ml A01AB81 Neomycin+ Benzocaine (dental) 30200 pieces A01AB82 Demeclocycline+ Triamcinolone (dental) 680 g A01AC Corticosteroids for local oral treatment A01AC81 Dexamethasone+ Thymol (dental) 3094 ml A01AD Other agents for local oral treatment A01AD80 Lidocaine+ Cetylpyridinium chloride (gingival) 227150 g A01AD81 Lidocaine+ Cetrimide (O) 30900 g A01AD82 Choline salicylate (O) 864720 pieces A01AD83 Lidocaine+ Chamomille extract (O) 370080 g A01AD90 Lidocaine+ Paraformaldehyde (dental) 405 g A02 DRUGS FOR ACID RELATED DISORDERS 47,1312 A02A ANTACIDS 1,0133 Combinations and complexes of aluminium, calcium and A02AD 1,0133 magnesium compounds A02AD81 Aluminium hydroxide+ Magnesium hydroxide (O) 811120 pieces 10 pieces 0,1689 A02AD81 Aluminium hydroxide+ Magnesium hydroxide (O) 3101974 ml 50 ml 0,1292 A02AD83 Calcium carbonate+ Magnesium carbonate (O) 3434232 pieces 10 pieces 0,7152 DRUGS FOR PEPTIC ULCER AND GASTRO- A02B 46,1179 OESOPHAGEAL REFLUX DISEASE (GORD) A02BA H2-receptor antagonists 2,3855 A02BA02 Ranitidine (O) 340327,5 g 0,3 g 2,3624 A02BA02 Ranitidine (P) 3318,25 g 0,3 g 0,0230 A02BC Proton pump inhibitors 43,7324 A02BC01 Omeprazole -

Household Insects – Homeowners ` CAUTION: All Insecticides Are Toxic to Some Degree; Therefore, Care Should Be Exercised in Their Use
Household Insects – Homeowners ` CAUTION: All insecticides are toxic to some degree; therefore, care should be exercised in their use. The manufacturer’s directions on the label in the use of the material must be followed explicitly. Insect Threats Insecticides and Treatment* Remarks Ants Feed on foods and Baits (active ingredient and Remove food and clean up the area. Place (several may damage product): bait where ants occur or congregate. May species) clothing; may also sodium tetraborate decahydrate use several different baits at the same time sting, causing severe (Amdro Kills Ants Liquid Bait, Terro to discover one that ants will consume. reaction to some Liquid Ant Baits); Care should be taken not to contaminate people. hydramethylnon (Amdro Kills Ants foodstuffs. Also treat nests in yard. Follow Bait Stations and Stakes); label. orthoboric acid (Terro Perimeter Ant Bait); fipronil (Combat Max Ant Killing Bait Stations and Gel); abamectin (Raid Max Double Control Ant Baits, Raid Ant Baits III); dinotefuran (Hot Shot Ultra Clear Roach & Ant Gel Bait, Hot Shot Ultra Liquid Ant Bait); spinosad (Ortho Home Defense Liquid Ant Bait); thiamethoxam (Raid Precision Placement Ant Bait Gel) Crack and crevices: Follow label. prallethrin, esfenvalerate, pyrethrins, pyrethrum, permethrin, tetra- methrin, phenothrin, beta-cyfluthrin, cyfluthrin Indoor space: prallethrin, esfenvalerate, pyrethrins, pyrethrum, permethrin, tetramethrin, phenothrin, cyfluthrin, bifenthrin Outdoor barrier: prallethrin, esfenvalerate, permethrin, beta-cyfluthrin, cyfluthrin, bifenthrin, malathion, carbaryl Outdoor broadcast: hydramethylnon, pyriproxyfen, beta-cyfluthrin, esfenvalerate, bifenthrin, cyfluthrin, malathion, carbaryl *Labels on insecticides should state “material may be used in the household” and should be registered by the EPA for that purpose. Household Insects – Homeowners ` CAUTION: All insecticides are toxic to some degree; therefore, care should be exercised in their use. -

The Toxic Effect of Permethrin and Cypermethrin on Engorged Ixodes
Annals of Agricultural and Environmental Medicine 2014, Vol 21, No 2, 259–262 ORIGINAL ARTICLE www.aaem.pl The toxic effect of permethrin and cypermethrin on engorged Ixodes ricinus females Alicja Buczek1, Katarzyna Bartosik1, Paweł Kuczyński1,2 1 Chair and Department of Biology and Parasitology, Medical University, Lublin, Poland 2 Chair and Department of Rehabilitation and Orthopaedics, Medical University, Lublin, Poland Buczek A, Bartosik K, Kuczyński P. The toxic effect of permethrin and cypermethrin on engorged Ixodes ricinus females. Ann Agric Environ Med. 2014; 21(2): 259–262. doi: 10.5604/1232-1966.1108587 Abstract Introduction. Ixodes ricinus tick is of great medical and veterinary importance and has a wide range of geographical distribution. The study presents the effect of permethrin (Per) and cypermethrin (CM) on engorged I. ricinus females. Materials and method. The effect of perythroids studied on engorged I. ricinus females was assessed on the basis of the pre-oviposition and oviposition period. Remote effects of Per and CM application were assessed by investigation of the length and course of embryonic development and larval hatching from eggs laid by pyrethroid-treated females. Per (Copex WP) was used at doses of 0.78125–25.0 µg/1 specimen, and CM (Kordon 10WP) was applied at 0.3125–10.0 µg/1 specimen. Immediately after the feeding period, I. ricinus females were sprayed with 20 µl of a pyrethroid solution and kept at 28 °C and 75%RH. Results. The experiments demonstrated that CM exerted a stronger toxic effect on I. ricinus females than Per. The lowest doses of CM doubled the length of the pre-oviposition period while its highest doses prolonged the period nearly three times compared with the control.