New Zealand Fur Seal Research Project
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

56. Otariidae and Phocidae
FAUNA of AUSTRALIA 56. OTARIIDAE AND PHOCIDAE JUDITH E. KING 1 Australian Sea-lion–Neophoca cinerea [G. Ross] Southern Elephant Seal–Mirounga leonina [G. Ross] Ross Seal, with pup–Ommatophoca rossii [J. Libke] Australian Sea-lion–Neophoca cinerea [G. Ross] Weddell Seal–Leptonychotes weddellii [P. Shaughnessy] New Zealand Fur-seal–Arctocephalus forsteri [G. Ross] Crab-eater Seal–Lobodon carcinophagus [P. Shaughnessy] 56. OTARIIDAE AND PHOCIDAE DEFINITION AND GENERAL DESCRIPTION Pinnipeds are aquatic carnivores. They differ from other mammals in their streamlined shape, reduction of pinnae and adaptation of both fore and hind feet to form flippers. In the skull, the orbits are enlarged, the lacrimal bones are absent or indistinct and there are never more than three upper and two lower incisors. The cheek teeth are nearly homodont and some conditions of the ear that are very distinctive (Repenning 1972). Both superfamilies of pinnipeds, Phocoidea and Otarioidea, are represented in Australian waters by a number of species (Table 56.1). The various superfamilies and families may be distinguished by important and/or easily observed characters (Table 56.2). King (1983b) provided more detailed lists and references. These and other differences between the above two groups are not regarded as being of great significance, especially as an undoubted fur seal (Australian Fur-seal Arctocephalus pusillus) is as big as some of the sea lions and has some characters of the skull, teeth and behaviour which are rather more like sea lions (Repenning, Peterson & Hubbs 1971; Warneke & Shaughnessy 1985). The Phocoidea includes the single Family Phocidae – the ‘true seals’, distinguished from the Otariidae by the absence of a pinna and by the position of the hind flippers (Fig. -

Brucella Antibody Seroprevalence in Antarctic Seals (Arctocephalus Gazella, Leptonychotes Weddellii and Mirounga Leonina)
Vol. 105: 175–181, 2013 DISEASES OF AQUATIC ORGANISMS Published September 3 doi: 10.3354/dao02633 Dis Aquat Org Brucella antibody seroprevalence in Antarctic seals (Arctocephalus gazella, Leptonychotes weddellii and Mirounga leonina) Silje-Kristin Jensen1,2,*, Ingebjørg Helena Nymo1, Jaume Forcada3, Ailsa Hall2, Jacques Godfroid1 1Section for Arctic Veterinary Medicine, Norwegian School of Veterinary Science, Stakkevollveien 23, 9010 Tromsø, Norway; member of the Fram Centre - High North Research Centre for Climate and the Environment, 9296 Tromsø, Norway 2Sea Mammal Research Unit, Scottish Oceans Institute, University of St. Andrews, St. Andrews KY16 8LB, UK 3British Antarctic Survey, Natural Environment Research Council, High Cross, Madingley Road, Cambridge CB3 0ET, UK ABSTRACT: Brucellosis is a worldwide infectious zoonotic disease caused by Gram-negative bac- teria of the genus Brucella, and Brucella infections in marine mammals were first reported in 1994. A serosurvey investigating the presence of anti-Brucella antibodies in 3 Antarctic pinniped spe- cies was undertaken with a protein A/G indirect enzyme-linked immunosorbent assay (iELISA) and the Rose Bengal test (RBT). Serum samples from 33 Weddell seals Leptonychotes weddelli were analysed, and antibodies were detected in 8 individuals (24.2%) with the iELISA and in 21 (65.6%) with the RBT. We tested 48 southern elephant seal Mirounga leonina sera and detected antibodies in 2 animals (4.7%) with both the iELISA and the RBT. None of the 21 Antarctic fur seals Arctocephalus gazella was found positive. This is the first report of anti-Brucella antibodies in southern elephant seals. The potential impact of Brucella infection in pinnipeds in Antarctica is not known, but Brucella spp. -

The Evolution of Infanticide by Females in Mammals
bioRxiv preprint doi: https://doi.org/10.1101/405688; this version posted September 3, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. The evolution of infanticide by females in mammals Dieter Lukas1,2* & Elise Huchard1,3 1) Department of Zoology, University of Cambridge, Downing Street, CB2 3EJ Cambridge, U. K. 2) Department of Human Behaviour, Ecology, and Culture, MPI for Evolutionary Anthropology, Deutscher Platz 6, 04103 Leipzig, Germany 3) Institut des Sciences de L'Evolution de Montpellier, UMR 5554, CNRS, Université de Montpellier, Montpellier, France *) Author for correspondence: [email protected] ORCID: Dieter Lukas: 0000-0002-7141-3545; Elise Huchard: 0000-0002-6944-449X Keywords: social competition; sexual selection; phylogenetic comparison; sociality; kinship Abstract In most mammalian species, females regularly interact with kin, and it may thus be difficult to understand the evolution of some aggressive and harmful competitive behaviour among females, such as infanticide. Here, we investigate the evolutionary determinants of infanticide by females by combining a quantitative analysis of the taxonomic distribution of infanticide with a qualitative synthesis of the circumstances of infanticidal attacks in published reports. Our results show that female infanticide is widespread across mammals and varies in relation to social organization and life-history, being more frequent where females breed in group and invest much energy into reproduction. Specifically, female infanticide occurs where the proximity of conspecific offspring directly threatens the killer’s reproductive success by limiting access to critical resources for her dependent progeny, including food, shelters, care or a social position. -

Marine Mammal Taxonomy
Marine Mammal Taxonomy Kingdom: Animalia (Animals) Phylum: Chordata (Animals with notochords) Subphylum: Vertebrata (Vertebrates) Class: Mammalia (Mammals) Order: Cetacea (Cetaceans) Suborder: Mysticeti (Baleen Whales) Family: Balaenidae (Right Whales) Balaena mysticetus Bowhead whale Eubalaena australis Southern right whale Eubalaena glacialis North Atlantic right whale Eubalaena japonica North Pacific right whale Family: Neobalaenidae (Pygmy Right Whale) Caperea marginata Pygmy right whale Family: Eschrichtiidae (Grey Whale) Eschrichtius robustus Grey whale Family: Balaenopteridae (Rorquals) Balaenoptera acutorostrata Minke whale Balaenoptera bonaerensis Arctic Minke whale Balaenoptera borealis Sei whale Balaenoptera edeni Byrde’s whale Balaenoptera musculus Blue whale Balaenoptera physalus Fin whale Megaptera novaeangliae Humpback whale Order: Cetacea (Cetaceans) Suborder: Odontoceti (Toothed Whales) Family: Physeteridae (Sperm Whale) Physeter macrocephalus Sperm whale Family: Kogiidae (Pygmy and Dwarf Sperm Whales) Kogia breviceps Pygmy sperm whale Kogia sima Dwarf sperm whale DOLPHIN R ESEARCH C ENTER , 58901 Overseas Hwy, Grassy Key, FL 33050 (305) 289 -1121 www.dolphins.org Family: Platanistidae (South Asian River Dolphin) Platanista gangetica gangetica South Asian river dolphin (also known as Ganges and Indus river dolphins) Family: Iniidae (Amazon River Dolphin) Inia geoffrensis Amazon river dolphin (boto) Family: Lipotidae (Chinese River Dolphin) Lipotes vexillifer Chinese river dolphin (baiji) Family: Pontoporiidae (Franciscana) -

Zoo Keeper Information
ZOO KEEPER INFORMATION Auckland Zoo and its role in Conservation and Captive Breeding Programmes Revised by Kirsty Chalmers Registrar 2006 CONTENTS Introduction 3 Auckland Zoo vision, mission and strategic intent 4 The role of modern zoos 5 Issues with captive breeding programmes 6 Overcoming captive breeding problems 7 Assessing degrees of risk 8 IUCN threatened species categories 10 Trade in endangered species 12 CITES 12 The World Zoo and Aquarium Conservation Strategy 13 International Species Information System (ISIS) 15 Animal Records Keeping System (ARKS) 15 Auckland Zoo’s records 17 Identification of animals 17 What should go on daily reports? 18 Zoological Information Management System (ZIMS) 19 Studbooks and SPARKS 20 Species co-ordinators and taxon advisory groups 20 ARAZPA 21 Australasian Species Management Program (ASMP) 21 Animal transfers 22 Some useful acronyms 24 Some useful references 25 Appendices 26 Zoo Keeper Information 2006 2 INTRODUCTION The intention of this manual is to give a basic overview of the general operating environment of zoos, and some of Auckland Zoo’s internal procedures and external relationships, in particular those that have an impact on species management and husbandry. The manual is designed to be of benefit to all keepers, to offer a better understanding of the importance of captive animal husbandry and species management on a national and international level. Zoo Keeper Information 2006 3 AUCKLAND ZOO VISION Auckland Zoo will be globally acknowledged as an outstanding, progressive zoological park. AUCKLAND ZOO MISSION To focus the Zoo’s resources to benefit conservation and provide exciting visitor experiences which inspire and empower people to take positive action for wildlife and the environment. -

Behaviour of Lactating Steller Sea Lions (Eumetopias Jubatus) During the Breeding Season
Behaviour of Lactating Steller Sea Lions (Eumetopias jubatus) During the Breeding Season: A Comparison between a Declining and Stable Population in Alaska by Linda Leontine Milette B.Sc. (Biology), Simon Fraser University, B.C. A thesis submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in The Faculty of Graduate Studies DEPARTMENT OF ZOOLOGY July 1999 We accept this thesis as conforming to the required standard THE UNIVERSITY OF BRITISH COLUMBIA © Linda Leontine Milette, 1999 In presenting this thesis in partial fulfilment of the requirements for an advanced degree at the University of British Columbia, I agree that the Library shall make it freely available for reference and study. I further agree that permission for extensive copying of this thesis for scholarly purposes may be granted by the head of my department or by his or her representatives. It is understood that copying or publication of this thesis for financial gain shall not be allowed without my written permission. Department of The University of British Columbia Vancouver, Canada DE-6 (2/88) Milette: Behaviour of Lactating Steller Sea Lions ABSTRACT Female attendance patterns and activity budgets of Alaskan Steller sea lions (Eumetopias jubatus) were compared at two sites using scan sampling over two summer breeding seasons in 1994 and 1995 at Sugarloaf Island (a declining population) and Lowrie Island (a stable population). The goal was to document female behaviour and to determine whether there were behavioural differences between the two sites that were consistent with the hypothesis that Steller sea lions in the area of population decline were food- limited. -

April 1St- 3Rd, 2016 Savannah State University
April 1st- 3rd, 2016 Savannah State University Schedule Saturday, April 2nd 8:00 Registration 8:30 Poster Set-up 9:00 Opening remarks 9:15 Panel Discussion: Life After SEAMAMMS Kelli Edwards, Damon Gannon, Robin Perrtree, Steve Thornton 10:15 Break Session 1 Chair: Damon Gannon 10:45 Christina Toms et al. A report on the potential influence of a record-breaking flood event on bottlenose dolphin (Tursiops truncatus) populations in Pensacola Bay, Florida. 11:00 Madison Miketa et al. Climate change and responses to seagrass die-off in a top predator, the bottlenose dolphin. 11:15 Bette Rubin Blue Whale Vocalizations off the Scotian Shelf using PAM: Analysis and Management Implications. 11:30 Hannah Nylander-Asplin et al. Acoustic Monitoring of Bottlenose Dolphins (Tursiops truncatus) in the May River, South Carolina. 11:45 Rachael Randall et al. Prey availability assessment of the common bottlenose dolphin Tursiops truncatus in two locations near Savannah, Georgia. 12:00 Lunch Session 2 Chair: Brian Balmer 13:30 Carissa King and Quincy Gibson. Identification of bottlenose dolphin (Tursiops truncatus) critical habitat areas in the St. Johns River, Florida. 13:45 Seth Klepal et al. Why there needs to be a universal definition of residency for common bottlenose dolphins (Tursiops truncatus) that can be used across different study areas. 14:00 Jessica Thompson et al. Estimation of site fidelity for common bottlenose dolphins Tursiops truncatus in a complex estuarine system using the Robust Design and multistate analysis. 14:15 Daniela Silva et al. Who, Where, and When? Bottlenose Dolphin (Tursiops truncatus) Stock Structure off Northern South Carolina. -

Behaviour of Lactating Steller Sea Lions (Eumetopias Jubatus) During the Breeding Season: a Comparison Between a Declining and Stable Population in Alaska
Behaviour of Lactating Steller Sea Lions (Eumetopias jubatus) During the Breeding Season: A Comparison between a Declining and Stable Population in Alaska Linda Leontine Milette B.Sc. (Biology), Simon Fraser University, B.C. A thesis submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in The Faculty of Graduate Studies DEPARTMENT OF ZOOLOGY July 1999 We accept this thesis as conforming to the required standard THE UNIVERSITY OF BRITISH COLUMBIA © Linda Leontine Milette, 1999 Milette: Behaviour of Lactating Steller Sea Lions ABSTRACT Female attendance patterns and activity budgets of Alaskan Steller sea lions (Eumetopias jubatus) were compared at two sites using scan sampling over two summer breeding seasons in 1994 and 1995 at Sugarloaf Island (a declining population) and Lowrie Island (a stable population). The goal was to document female behaviour and to determine whether there were behavioural differences between the two sites that were consistent with the hypothesis that Steller sea lions in the area of population decline were food- limited. The perinatal period (time from birth of pup to the mother’s first feeding trip) averaged 10.1 days in the area of population decline compared to 8.0 days in the stable area, counter to initial predictions. The first shore visit following the perinatal period was significantly longer in the area of population decline compared to the stable population. Females from both populations exhibited a diel haul out pattern where the majority of returns and departures to and from the rookery occurred between 1800 - 0600 hours. Similarly, the mean length of female foraging trips at both populations increased as their pups grew older, whereas shore visits became shorter. -

List of 28 Orders, 129 Families, 598 Genera and 1121 Species in Mammal Images Library 31 December 2013
What the American Society of Mammalogists has in the images library LIST OF 28 ORDERS, 129 FAMILIES, 598 GENERA AND 1121 SPECIES IN MAMMAL IMAGES LIBRARY 31 DECEMBER 2013 AFROSORICIDA (5 genera, 5 species) – golden moles and tenrecs CHRYSOCHLORIDAE - golden moles Chrysospalax villosus - Rough-haired Golden Mole TENRECIDAE - tenrecs 1. Echinops telfairi - Lesser Hedgehog Tenrec 2. Hemicentetes semispinosus – Lowland Streaked Tenrec 3. Microgale dobsoni - Dobson’s Shrew Tenrec 4. Tenrec ecaudatus – Tailless Tenrec ARTIODACTYLA (83 genera, 142 species) – paraxonic (mostly even-toed) ungulates ANTILOCAPRIDAE - pronghorns Antilocapra americana - Pronghorn BOVIDAE (46 genera) - cattle, sheep, goats, and antelopes 1. Addax nasomaculatus - Addax 2. Aepyceros melampus - Impala 3. Alcelaphus buselaphus - Hartebeest 4. Alcelaphus caama – Red Hartebeest 5. Ammotragus lervia - Barbary Sheep 6. Antidorcas marsupialis - Springbok 7. Antilope cervicapra – Blackbuck 8. Beatragus hunter – Hunter’s Hartebeest 9. Bison bison - American Bison 10. Bison bonasus - European Bison 11. Bos frontalis - Gaur 12. Bos javanicus - Banteng 13. Bos taurus -Auroch 14. Boselaphus tragocamelus - Nilgai 15. Bubalus bubalis - Water Buffalo 16. Bubalus depressicornis - Anoa 17. Bubalus quarlesi - Mountain Anoa 18. Budorcas taxicolor - Takin 19. Capra caucasica - Tur 20. Capra falconeri - Markhor 21. Capra hircus - Goat 22. Capra nubiana – Nubian Ibex 23. Capra pyrenaica – Spanish Ibex 24. Capricornis crispus – Japanese Serow 25. Cephalophus jentinki - Jentink's Duiker 26. Cephalophus natalensis – Red Duiker 1 What the American Society of Mammalogists has in the images library 27. Cephalophus niger – Black Duiker 28. Cephalophus rufilatus – Red-flanked Duiker 29. Cephalophus silvicultor - Yellow-backed Duiker 30. Cephalophus zebra - Zebra Duiker 31. Connochaetes gnou - Black Wildebeest 32. Connochaetes taurinus - Blue Wildebeest 33. Damaliscus korrigum – Topi 34. -

Female Reproductive Senescence Across
Female reproductive senescence across mammals: A high diversity of patterns modulated by life history and mating traits Jean-François Lemaître, Victor Ronget, Jean-Michel Gaillard To cite this version: Jean-François Lemaître, Victor Ronget, Jean-Michel Gaillard. Female reproductive senescence across mammals: A high diversity of patterns modulated by life history and mating traits. Mechanisms of Ageing and Development, Elsevier, 2020, 192, pp.111377. 10.1016/j.mad.2020.111377. hal-03060282 HAL Id: hal-03060282 https://hal.archives-ouvertes.fr/hal-03060282 Submitted on 14 Dec 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. 1 Female reproductive senescence 2 across mammals: a high diversity of 3 patterns modulated by life history 4 and mating traits 5 6 Jean-François Lemaître1, Victor Ronget2 & Jean-Michel Gaillard1 7 8 9 1 Univ Lyon, Université Lyon 1, CNRS, Laboratoire de Biométrie et Biologie Évolutive UMR 5558, F-69622, 10 Villeurbanne, France. 11 2 Unité Eco-anthropologie (EA), Muséum National d’Histoire Naturelle, CNRS, Université Paris Diderot, F- 12 75016 Paris, France. 13 14 15 16 Article for the special issue “Understanding the biology of aging to better intervene” 17 18 19 20 1 21 ABSTRACT 22 23 Senescence patterns are highly variable across the animal kingdom. -
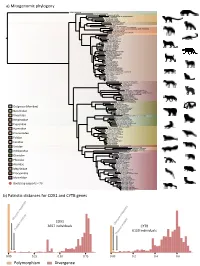
A) Mitogenomic Phylogeny B) Patristic Distances for COX1 and CYTB
a) Mitogenomic phylogeny Manis javanica Nandinia binotata Paguma larvata Paradoxurus hermaphroditus Genetta servalina Viverricula indica Civettictis civetta Viverra tangalunga Bdeogale nigripes Urva javanica Fossa fossana Mungotictis decemlineata Proteles cristata cristataTS307 Proteles cristata septentrionalis NMS Z 2018 54 Crocuta crocuta Hyaena hyaena Parahyaena brunnea FD126 Prionodon pardicolor Neofelis nebulosa Panthera tigris Panthera onca Panthera uncia Panthera leo Panthera pardus Leopardus tigrinus Leopardus geoffroyi Leopardus guigna Leopardus colocolo Leopardus jacobita Leopardus pardalis Leopardus wiedii Pardofelis marmorata Catopuma badia Catopuma temminckii Acinonyx jubatus Puma concolor Puma yagouaroundi Lynx rufus Lynx lynx Lynx canadensis Lynx pardinus Leptailurus serval Caracal caracal Profelis aurata Prionailurus rubiginosus Prionailurus planiceps Prionailurus bengalensis Prionailurus bengalensis euptilurus Prionailurus viverrinus Otocolobus manul Felis chaus Felis nigripes Felis margarita Felis catus Felis silvestris Chrysocyon brachyurus Speothos venaticus Lycaon pictus Cuon alpinus Canis latrans Canis anthus Canis lupus Urocyon cinereoargenteus Otocyon megalotis megalotis TS305 Otocyon megalotis virgatus FMNH158128 Nyctereutes procyonoides Vulpes lagopus Vulpes corsac Vulpes ferrilata Vulpes vulpes T3611 Vulpes zerda Ailuropoda melanoleuca Arctodus simus Arctotherium sp Tremarctos ornatus Outgroup (Manidae) Melursus ursinus Helarctos malayanus Ursus americanus Ursus thibetanus Nandinidae Ursus spelaeus Ursus arctos -

Fur Seals and Sea Lions (Otariidae): Identification of Species and Taxonomic Review
Systematics and Biodiversity 1 (3): 339–439 Issued 16 February 2004 DOI: 10.1017/S147720000300121X Printed in the United Kingdom C The Natural History Museum Fur seals and sea lions (Otariidae): identification of species and taxonomic review Sylvia Brunner Australian Marine Mammal Research Centre, University of Sydney, NSW 2006, Australia CSIRO Sustainable Ecosystems, GPO Box 284, Canberra City, ACT 2601, Australia ∗ University of Alaska Museum, 907 Yukon Drive, Fairbanks AK 99775, United States submitted December 2002 accepted May 2003 Contents Abstract 340 Introduction 340 Materials and methods 343 Results 344 Description of species: sea lions Steller sea lion – Eumetopias jubatus 345 Southern sea lion – Otaria byronia 352 Australian sea lion – Neophoca cinerea 354 Hooker’s sea lion – Phocarctos hookeri 354 California sea lion – Zalophus californianus californianus 357 Galapagos sea lion – Zalophus californianus wollebaeki 357 Japanese sea lion – Zalophus californianus japonicus 360 Description of species: fur seals Northern fur seal – Callorhinus ursinus 360 Antarctic fur seal – Arctocephalus gazella 363 Subantarctic fur seal – Arctocephalus tropicalis 363 New Zealand fur seal – Arctocephalus forsteri 366 South African fur seal – Arctocephalus pusillus pusillus 366 Australian fur seal – Arctocephalus pusillus doriferus 368 Guadalupe fur seal – Arctocephalus townsendi 370 Galapagos fur seal – Arctocephalus galapagoensis 370 South American fur seal – Arctocephalus australis 371 Juan Fernandez fur seal – Arctocephalus philippii 374 A comparison of subspecies Arctocephalus pusillus 374 Zalophus californianus 374 Arctocephalus australis 375 The Otariidae 377 Discussion 378 References 384 Appendices I Summary details of specimens 386 II Univariate statistics for male and female otariids 405 ∗Correspondence address 339 340 Sylvia Brunner Abstract The standard anatomical descriptions given to identify species of the family Otariidae (fur seals and sea lions), particularly those for the genus Arctocephalus, have been largely inconclusive.