Fermentation of Dihydroxyacetone by Engineered Escherichia Coli and Klebsiella Variicola to Products
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Inhibition of Neuroinflammatory Nitric Oxide Signaling Suppresses Glycation and Prevents Neuronal Dysfunction in Mouse Prion Disease
Inhibition of neuroinflammatory nitric oxide signaling suppresses glycation and prevents neuronal dysfunction in mouse prion disease Julie-Myrtille Bourgognona, Jereme G. Spiersb, Sue W. Robinsonc, Hannah Scheiblichd, Paul Glynnc, Catharine Ortorie, Sophie J. Bradleyf, Andrew B. Tobinf, and Joern R. Steinertg,1 aCentre for Immunobiology, University of Glasgow, Glasgow, G12 8TA, United Kingdom; bDepartment of Biochemistry and Genetics, La Trobe Institute for Molecular Science, La Trobe University, VIC 3083, Melbourne, Australia; cMedical Research Council Toxicology Unit, University of Leicester, Leicester, LE1 9HN, United Kingdom; dDepartment of Neurodegenerative Disease and Geriatric Psychiatry/Neurology, University of Bonn, Bonn 53127, Germany; eSchool of Pharmacy, University of Nottingham, Nottingham NG7 2RD, United Kingdom; fCentre for Translational Pharmacology, Veterinary and Life Sciences, University of Glasgow, Glasgow, G12 8QQ, United Kingdom; and gSchool of Life Sciences, Queen’s Medical Centre, University of Nottingham, Nottingham, NG7 2UH, United Kingdom Edited by Juan Carlos Saez, University of Valparaiso, Valparaiso, Chile, and approved January 22, 2021 (received for review May 14, 2020) Several neurodegenerative diseases associated with protein mis- system where it is synthesized by NO synthases (neuronal NOS folding (Alzheimer’s and Parkinson’s disease) exhibit oxidative and [nNOS], inducible NOS [iNOS], and endothelial NOS [eNOS]). nitrergic stress following initiation of neuroinflammatory path- Excessive NO production -

Nomination Background: Dihydroxyacetone (CASRN: 96-26-4)
SUMMARY OF DATA FOR CHEMICAL SELECTION Dihydroxyacetone 96-26-4 BASIS OF NOMINATION TO THE CSWG As consumers have become more mindful of the hazards ofa "healthy tan," more individuals have turned to sunless tanning. Sunless tanning products represent about 10% of the $400 million market for suntan preparations, and these products are the fastest growing segment of the suntanning preparation market. All sunless tanners contain dihydroxyacetone. Information on the toxicity of dihydroxyacetone appears contradictory. A mutagen that induces DNA strand breaks, dihydroxyacetone is also an intermediate in carbohydrate metabolism in higher plants and animals. Such contradictions are not unprecedented, and it has been suggested that autooxidation of cx-hydroxycarbonyl compounds including reducing sugars may play a role in diseases associated with age and diabetes (Morita, 1991 ). When dihydroxyacetone was applied to the skin of mice, no carcinogenic effect was observed. It is unclear whether this negative response was caused by a failure ofthe compound to penetrate the skin. If so, extrapolating the dermal results to other routes of exposure would not be appropriate. NCI is nominating dihydroxyacetone to the NTP for dermal penetration studies in rats and mice to determine whether dihydroxyacetone can penetrate the skin. This information will clarify whether additional testing of dihydroxyacetone is warranted. Dihydroxyacetone 96-26-4 CHEMICAL IDENTIFICATION CAS Registry Number: 96-26-4 Chemical Abstracts Service Name: 1,3-Dihydroxy-2-propanone (9CI; 8CI) Synonyms and Tradenames: 1,3-Dihydroxydimethyl ketone; Chromelin; CTF A 00816; Dihyxal; Otan; Oxantin; Oxatone; Soleal; Triulose; Viticolor Structural Class: Ketone, ketotriose compound Structure. Molecular Formula. and Molecular Weight: 0 II /c"-. -

The Pichia Pastoris Dihydroxyacetone Kinase Is a PTS1-Containing, but Cytosolic, Protein That Is Essential for Growth on Methanol
. 14: 759–771 (1998) The Pichia pastoris Dihydroxyacetone Kinase is a PTS1-containing, but Cytosolic, Protein that is Essential for Growth on Methanol GEORG H. LU} ERS†, RAJ ADVANI, THIBAUT WENZEL AND SURESH SUBRAMANI* Department of Biology, University of California at San Diego, La Jolla, California 92093–0322, U.S.A. Received 11 November 1997; accepted 27 January 1998 Dihydroxyacetone kinase (DAK) is essential for methanol assimilation in methylotrophic yeasts. We have cloned the DAK gene from Pichia pastoris by functional complementation of a mutant that was unable to grow on methanol. An open reading frame of 1824 bp was identified that encodes a 65·3 kDa protein with high homology to DAK from Saccharomyces cerevisiae. Although DAK from P. pastoris contained a C-terminal tripeptide, TKL, which we showed can act as a peroxisomal targeting signal when fused to the green fluorescent protein, the enzyme was primarily cytosolic. The TKL tripeptide was not required for the biochemical function of DAK because a deletion construct lacking the DNA encoding this tripeptide was able to complement the P. pastoris dakÄ mutant. Peroxisomes, which are essential for growth of P. pastoris on methanol, were present in the dakÄ mutant and the import of peroxisomal proteins was not disturbed. The dakÄ mutant grew at normal rates on glycerol and oleate media. However, unlike the wild-type cells, the dakÄ mutant was unable to grow on methanol as the sole carbon source but was able to grow on dihydroxyacetone at a much slower rate. The metabolic pathway explaining the reduced growth rate of the dakÄ mutant on dihydroxyacetone is discussed. -

Matrix Scientific PO BOX 25067 COLUMBIA, SC 29224-5067 Telephone: 803-788-9494 Fax: 803-788-9419 SAFETY DATA SHEET Transportation Emergency: 3E Co
Matrix Scientific PO BOX 25067 COLUMBIA, SC 29224-5067 Telephone: 803-788-9494 Fax: 803-788-9419 SAFETY DATA SHEET Transportation Emergency: 3E Co. (5025) 800-451-8346 1. Product Identification Name 1,3-Dihydroxyacetone Catalog Number 119322 CAS Registry Number [96-26-4] Company Matrix Scientific Physical Address 131 Pontiac Business Center Drive Elgin, SC 29045 USA Telephone/Fax (803)788-9494/(803)788-9419 2. Hazard Identification Hazardous Ingredients 1,3-Dihydroxyacetone GHS label elements, including precautionary statements Pictogram Signal word WARNING Hazard statement(s) H317 H317 May cause an allergic skin reaction H319 H319 Causes serious eye irritation Precautionary statement(s) P280 Wear protective gloves/protective clothing/eye protection/face protection. P305+351+338 IF IN EYES: Rinse cautiously with water for several minutes. Remove contact lenses if present and easy to do - continue rinsing. P411 Store at temperatures not exceeding 0°C 3. Composition, Information or Ingredients Name 1,3-Dihydroxyacetone 4. First Aid Measures 1 Last Updated 11/20/2018 Eye Contact: Check for and remove any contact lenses. Immediately flush eyes with clean, running water for at least 15 minutes while keeping eyes open. Cool water may be used. Seek medical attention. Skin Contact: After contact with skin, wash with generous quantities of running water. Gently and thoroughly wash affected area with running water and non- abrasive soap. Cool water may be used. Cover the affected area with emollient. Seek medical attention. Wash any contaminated clothing prior to reusing. Inhalation: Remove the victim from the source of exposure to fresh, uncontaminated air. If victim's breathing is difficult, administer oxygen. -

Acetone Thermally Treated in Different Solvents Joji Okumura, Tetsuya Yanai, Izumi Yajima and Kazuo Hayashi Kawasaki Research Center, T
Agric. Biol. Chem., 54 (7), 1631-1638, 1990 1631 Volatile Products Formed from L-Cysteine and Dihydroxy- acetone Thermally Treated in Different Solvents Joji Okumura, Tetsuya Yanai, Izumi Yajima and Kazuo Hayashi Kawasaki Research Center, T. Hasegawa Co., Ltd., 335 Kariyado, Nakahara-ku, Kawasaki 211, Japan Received November 21, 1989 Equimolecular amounts of L-cysteine and dihydroxyacetone were heated at 110°C for 3hr in different solvent systems such as deionized water, glycerine, or triglyceride. The resulting mixtures were vacuum steam-distilled and each distillate was extracted with ethyl ether. The volatiles in the ether extracts were analyzed by gas chromatographyand gas chromatograph\-massspectrometry. Differences in the quality and quantity of volatiles formed in the systems were observed. Pyrazines, thiazoles, thiophenes, and someother sulfur-containing compounds wereidentified in the volatiles. Dimethylpyrazines were formed as major volatiles in the glycerine and triglyceride systems but were minor in the water system. 2-Acetylthiazole in the triglyceride system and 2-acetylthiophene in the glycerine system were secondary abundant products. In the water system, l-mercapto-2-propanone was found as a major volatile compound, and thiophenes as the next dominants. The Maillard reaction is significant in the sugar alcohols, oils and fats, or their mixtures formation of flavors from various heated were used for various applications. It has been foods. In the flavor industry, the Maillard observed in some cases that the quality of reaction -

Influence of Battery Power Setting on Carbonyl Emissions from Electronic Cigarettes
Tobacco Induced Diseases Short Report Influence of battery power setting on carbonyl emissions from electronic cigarettes Zuzana Zelinkova1, Thomas Wenzl1 ABSTRACT INTRODUCTION Although e-cigarettes share common features such as power units, heating elements and e-liquids, the variability in design and possibility for AFFILIATION customization represent potential risks for consumers. A main health concern is 1 Joint Research Centre, European Commission, Geel, the exposure to carbonyl compounds, which are formed from the main components Belgium of e-liquids, propylene glycol and glycerol, through thermal decomposition. Levels CORRESPONDENCE TO of carbonyl emissions in e-cigarette aerosols depend, amongst others, on the Thomas Wenzl. Joint Research power supplied to the coil. Thus, e-cigarettes with adjustable power outputs Centre, European Commission, Retieseweg 111, B-2440 Geel, might lead to high exposures to carbonyls if the users increase the power output Belgium. E-mail: Thomas. excessively. The aim of this work was to elucidate the generation of carbonyls in [email protected] relation to undue battery power setting. ORCID ID: https://orcid. org/0000-0003-2017-3788 METHODS Carbonyl emissions were generated by two modular e-cigarettes equipped with two atomizers containing coils of different resistance following the ISO KEYWORDS emission, electronic 20768:2018 method. The battery power output was increased from the lower cigarettes, vaping, carbonyls, wattage level to above the power range recommended by the producer. Carbonyls power setting were trapped by a 2,4-dinitrophenylhydrazine (DNPH) solution and analysed by Received: 25 June 2020 LC-MS/MS. Revised: 22 July 2020 Accepted: 14 August 2020 RESULTS The amount of carbonyl emissions increased with increasing power setting. -

Electron Transport Phosphorylation in Rumen Butyrivibrios: Unprecedented ATP Yield for Glucose Fermentation to Butyrate
HYPOTHESIS AND THEORY published: 24 June 2015 doi: 10.3389/fmicb.2015.00622 Electron transport phosphorylation in rumen butyrivibrios: unprecedented ATP yield for glucose fermentation to butyrate Timothy J. Hackmann1 and Jeffrey L. Firkins2* 1 Department of Animal Sciences, University of Florida, Gainesville, FL, USA, 2 Department of Animal Sciences, The Ohio State University, Columbus, OH, USA From a genomic analysis of rumen butyrivibrios (Butyrivibrio and Pseudobutyrivibrio sp.), we have re-evaluated the contribution of electron transport phosphorylation (ETP) to ATP formation in this group. This group is unique in that most (76%) genomes were predicted to possess genes for both Ech and Rnf transmembrane ion pumps. These pumps act in concert with the NifJ and Bcd-Etf to form a electrochemical potential (μH+ and μNa+), which drives ATP synthesis by ETP. Of the 62 total butyrivibrio genomes currently available from the Hungate 1000 project, all 62 were predicted to Edited by: possess NifJ, which reduces oxidized ferredoxin (Fdox) during pyruvate conversion to Emilio M. Ungerfeld, Instituto de Investigaciones acetyl-CoA. All 62 possessed all subunits of Bcd-Etf, which reduces Fdox and oxidizes Agropecuarias, Chile reduced NAD during crotonyl-CoA reduction. Additionally, 61 genomes possessed all Reviewed by: subunits of the Rnf, which generates μH+ or μNa+ from oxidation of reduced Fd Wolfgang Buckel, (Fdred) and reduction of oxidized NAD. Further, 47 genomes possessed all six subunits Philipps-Universität Marburg, + Germany of the Ech, which generates μH from oxidation of Fdred. For glucose fermentation Robert J. Wallace, to butyrate and H2, the electrochemical potential established should drive synthesis University of Aberdeen, UK of ∼1.5 ATP by the F0F1-ATP synthase (possessed by all 62 genomes). -

Valpos As Efficient Catalysts for Glycerol Conversion to Methanol
catalysts Article VAlPOs as Efficient Catalysts for Glycerol Conversion to Methanol 1, , 2, 2 2 Gheorghit, a Mitran * y, Florentina Neat, u y,S, tefan Neat, u , Mihaela M. Trandafir and Mihaela Florea 1,2,* 1 Department of Organic Chemistry, University of Bucharest, Biochemistry & Catalysis, Faculty of Chemistry, 4-12, Blv. Regina Elisabeta, 030018 Bucharest, Romania 2 National Institute of Material Physics, 405A Atomi¸stilor, P.O. Box MG 7, 077125 Măgurele, Romania; florentina.neatu@infim.ro (F.N.); stefan.neatu@infim.ro (S, .N.); mihaela.trandafir@infim.ro (M.M.T.) * Correspondence: [email protected] (G.M.); mihaela.florea@infim.ro (M.F.) Authors with equal contribution. y Received: 24 April 2020; Accepted: 28 June 2020; Published: 1 July 2020 Abstract: The catalytic activity of a series of vanadium aluminophosphates catalysts prepared by sol-gel method followed by combustion of the obtained gel was evaluated in glycerol conversion towards methanol. The materials were characterized by several techniques such as X-ray diffraction (XRD), UV-vis, Fourier-transform infrared (FTIR), Raman and X-ray photoelectron (XPS) spectroscopies. The amount of vanadium incorporated in aluminophosphates framework played an important role in the catalytic activity, while in the products distribution the key role is played by the vanadium oxidation state on the surface. The sample that contains a large amount of V4+ has the highest selectivity towards methanol. On the sample with the lowest vanadium loading the oxidation path to dihydroxyacetone is predominant. The catalyst with higher content of tetrahedral isolated vanadium species, such V5APO, is less active in breaking the C–C bonds in the glycerol molecule than the one containing polymeric species. -

Supplementary Informations SI2. Supplementary Table 1
Supplementary Informations SI2. Supplementary Table 1. M9, soil, and rhizosphere media composition. LB in Compound Name Exchange Reaction LB in soil LBin M9 rhizosphere H2O EX_cpd00001_e0 -15 -15 -10 O2 EX_cpd00007_e0 -15 -15 -10 Phosphate EX_cpd00009_e0 -15 -15 -10 CO2 EX_cpd00011_e0 -15 -15 0 Ammonia EX_cpd00013_e0 -7.5 -7.5 -10 L-glutamate EX_cpd00023_e0 0 -0.0283302 0 D-glucose EX_cpd00027_e0 -0.61972444 -0.04098397 0 Mn2 EX_cpd00030_e0 -15 -15 -10 Glycine EX_cpd00033_e0 -0.0068175 -0.00693094 0 Zn2 EX_cpd00034_e0 -15 -15 -10 L-alanine EX_cpd00035_e0 -0.02780553 -0.00823049 0 Succinate EX_cpd00036_e0 -0.0056245 -0.12240603 0 L-lysine EX_cpd00039_e0 0 -10 0 L-aspartate EX_cpd00041_e0 0 -0.03205557 0 Sulfate EX_cpd00048_e0 -15 -15 -10 L-arginine EX_cpd00051_e0 -0.0068175 -0.00948672 0 L-serine EX_cpd00054_e0 0 -0.01004986 0 Cu2+ EX_cpd00058_e0 -15 -15 -10 Ca2+ EX_cpd00063_e0 -15 -100 -10 L-ornithine EX_cpd00064_e0 -0.0068175 -0.00831712 0 H+ EX_cpd00067_e0 -15 -15 -10 L-tyrosine EX_cpd00069_e0 -0.0068175 -0.00233919 0 Sucrose EX_cpd00076_e0 0 -0.02049199 0 L-cysteine EX_cpd00084_e0 -0.0068175 0 0 Cl- EX_cpd00099_e0 -15 -15 -10 Glycerol EX_cpd00100_e0 0 0 -10 Biotin EX_cpd00104_e0 -15 -15 0 D-ribose EX_cpd00105_e0 -0.01862144 0 0 L-leucine EX_cpd00107_e0 -0.03596182 -0.00303228 0 D-galactose EX_cpd00108_e0 -0.25290619 -0.18317325 0 L-histidine EX_cpd00119_e0 -0.0068175 -0.00506825 0 L-proline EX_cpd00129_e0 -0.01102953 0 0 L-malate EX_cpd00130_e0 -0.03649016 -0.79413596 0 D-mannose EX_cpd00138_e0 -0.2540567 -0.05436649 0 Co2 EX_cpd00149_e0 -
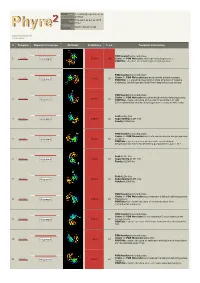
Phyre 2 Results for P25553
Email [email protected] Description P25553 Thu Jan 5 11:42:11 GMT Date 2012 Unique Job 5024f4b9e5342484 ID Detailed template information # Template Alignment Coverage 3D Model Confidence % i.d. Template Information PDB header:oxidoreductase 1 c2hg2A_ Alignment 100.0 100 Chain: A: PDB Molecule:aldehyde dehydrogenase a; PDBTitle: structure of lactaldehyde dehydrogenase PDB header:oxidoreductase Chain: B: PDB Molecule:betaine aldehyde dehydrogenase; 2 c3ed6B_ 100.0 36 Alignment PDBTitle: 1.7 angstrom resolution crystal structure of betaine aldehyde2 dehydrogenase (betb) from staphylococcus aureus PDB header:oxidoreductase Chain: A: PDB Molecule:formyltetrahydrofolate dehydrogenase; 3 c2o2qA_ 100.0 33 Alignment PDBTitle: crystal structure of the c-terminal domain of rat2 10'formyltetrahydrofolate dehydrogenase in complex with nadp Fold:ALDH-like 4 d1a4sa_ Alignment 100.0 35 Superfamily:ALDH-like Family:ALDH-like PDB header:oxidoreductase Chain: H: PDB Molecule:succinate-semialdehyde dehydrogenase 5 c3ifgH_ Alignment 100.0 34 (nadp+); PDBTitle: crystal structure of succinate-semialdehyde dehydrogenase from2 burkholderia pseudomallei, part 1 of 2 Fold:ALDH-like 6 d1bxsa_ Alignment 100.0 33 Superfamily:ALDH-like Family:ALDH-like Fold:ALDH-like 7 d1o9ja_ Alignment 100.0 33 Superfamily:ALDH-like Family:ALDH-like PDB header:oxidoreductase Chain: A: PDB Molecule:succinate-semialdehyde dehydrogenase 8 c3rh9A_ Alignment 100.0 35 (nad(p)(+)); PDBTitle: the crystal structure of oxidoreductase from marinobacter aquaeolei PDB header:oxidoreductase Chain: B: PDB Molecule:5-carboxymethyl-2-hydroxymuconate 9 c2d4eB_ Alignment 100.0 32 semialdehyde PDBTitle: crystal structure of the hpcc from thermus thermophilus hb8 PDB header:oxidoreductase Chain: G: PDB Molecule:antiquitin; 10 c2jg7G_ 100.0 28 Alignment PDBTitle: crystal structure of seabream antiquitin and elucidation of2 its substrate specificity PDB header:oxidoreductase Chain: C: PDB Molecule:succinate-semialdehyde dehydrogenase 11 c3jz4C_ 100.0 39 Alignment [nadp+]; PDBTitle: crystal structure of e. -

Fermentation of Dihydroxyacetone by Engineered Escherichia Coli and Klebsiella Variicola to Products
Fermentation of dihydroxyacetone by engineered Escherichia coli and Klebsiella variicola to products Liang Wanga, Diane Chauliaca,1, Mun Su Rheea,2, Anushadevi Panneerselvama, Lonnie O. Ingrama,3, and K. T. Shanmugama,3 aDepartment of Microbiology and Cell Science, University of Florida, Gainesville, FL 32611 Contributed by Lonnie O. Ingram, March 21, 2018 (sent for review January 18, 2018; reviewed by John W. Frost and F. Robert Tabita) Methane can be converted to triose dihydroxyacetone (DHA) by process, formaldehyde can also be produced biologically from chemical processes with formaldehyde as an intermediate. Carbon CO2 with formate as an intermediate (Fig. 1) (7). Dickens and dioxide, a by-product of various industries including ethanol/ Williamson reported as early as 1958 that DHA can be produced butanol biorefineries, can also be converted to formaldehyde biologically by transketolation of hydroxypyruvate and formalde- and then to DHA. DHA, upon entry into a cell and phosphorylation hyde (8). This transketolase is implicated in a unique pentose– to DHA-3-phosphate, enters the glycolytic pathway and can be phosphate–dependent pathway (DHA cycle) in methanol-utilizing fermented to any one of several products. However, DHA is yeast that fixes formaldehyde to xylulose-5-phosphate, yielding inhibitory to microbes due to its chemical interaction with cellular DHA as an intermediate in the production of glyceraldehyde-3- components. Fermentation of DHA to D-lactate by Escherichia coli phosphate in a cyclic mode (9). DHA in the cytoplasm is phos- strain TG113 was inefficient, and growth was inhibited by 30 g·L−1 phorylated by DHA kinase and/or glycerol kinase, and the DHA-P DHA. -

Table 4. V. Cholerae Flexgene ORF Collection
Table 4. V. cholerae FLEXGene ORF collection Reference Clone protein PlasmID clone GenBank Locus tag Symbol accession identifier FLEX clone name accession Product name VC0001 NP_062585 VcCD00019918 FLH200476.01F DQ772770 hypothetical protein VC0002 mioC NP_062586 VcCD00019938 FLH200506.01F DQ772771 mioC protein VC0003 thdF NP_062587 VcCD00019958 FLH200531.01F DQ772772 thiophene and furan oxidation protein ThdF VC0004 yidC NP_062588 VcCD00019970 FLH200545.01F DQ772773 inner membrane protein, 60 kDa VC0005 NP_062589 VcCD00061243 FLH236482.01F DQ899316 conserved hypothetical protein VC0006 rnpA NP_062590 VcCD00025697 FLH214799.01F DQ772774 ribonuclease P protein component VC0007 rpmH NP_062591 VcCD00061229 FLH236450.01F DQ899317 ribosomal protein L34 VC0008 NP_062592 VcCD00019917 FLH200475.01F DQ772775 amino acid ABC transporter, ATP-binding protein VC0009 NP_062593 VcCD00019966 FLH200540.01F DQ772776 amino acid ABC transproter, permease protein VC0010 NP_062594 VcCD00019152 FLH199275.01F DQ772777 amino acid ABC transporter, periplasmic amino acid-binding portion VC0011 NP_062595 VcCD00019151 FLH199274.01F DQ772778 hypothetical protein VC0012 dnaA NP_062596 VcCD00017363 FLH174286.01F DQ772779 chromosomal DNA replication initiator DnaA VC0013 dnaN NP_062597 VcCD00017316 FLH174063.01F DQ772780 DNA polymerase III, beta chain VC0014 recF NP_062598 VcCD00019182 FLH199319.01F DQ772781 recF protein VC0015 gyrB NP_062599 VcCD00025458 FLH174642.01F DQ772782 DNA gyrase, subunit B VC0016 NP_229675 VcCD00019198 FLH199346.01F DQ772783 hypothetical protein