Management of Arthropod Pathogen Vectors in North America: Minimizing Adverse Effects on Pollinators
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Foraging Behavior of Anax Junius (Odonata: Aeschnidae) and Its Potential As a Behavioral Endpoint in Pesticide Testing Sandra Kaye Brewer Iowa State University
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1997 The foraging behavior of Anax junius (Odonata: Aeschnidae) and its potential as a behavioral endpoint in pesticide testing Sandra Kaye Brewer Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Ecology and Evolutionary Biology Commons, Entomology Commons, Environmental Sciences Commons, Fresh Water Studies Commons, Toxicology Commons, and the Zoology Commons Recommended Citation Brewer, Sandra Kaye, "The foraging behavior of Anax junius (Odonata: Aeschnidae) and its potential as a behavioral endpoint in pesticide testing " (1997). Retrospective Theses and Dissertations. 11445. https://lib.dr.iastate.edu/rtd/11445 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. UMI MICROFILMED 1997 INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter fiice, while others may be from any type of computer printer. The quality of this reprcduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. -

Notes on Southern Africa Jerusalem Crickets (Orthoptera: Stenopelmatidae: Sia)
Zootaxa 3616 (1): 049–060 ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2013 Magnolia Press ISSN 1175-5334 (online edition) http://dx.doi.org/10.11646/zootaxa.3616.1.4 http://zoobank.org/urn:lsid:zoobank.org:pub:C790EC39-8BE6-475A-9CA8-3FBC443D3F93 Notes on southern Africa Jerusalem crickets (Orthoptera: Stenopelmatidae: Sia) DAVID B. WEISSMAN1* & CORINNA S. BAZELET 2,3 1Department of Entomology, California Academy of Sciences, San Francisco, CA, 94118, U.S.A. 2Steinhardt Museum, Department of Zoology, Faculty of Life Sciences, Tel Aviv University, Tel Aviv 69978, ISRAEL. 3Department of Conservation Ecology and Entomology, Stellenbosch University, Private Bag X1, Matieland 7602, SOUTH AFRICA. *To whom correspondence should be addressed. E-mail: [email protected] Abstract The Old World Jerusalem cricket (JC) subfamily Siinae contains one genus, Sia, with two subgenera: Sia (Sia) with two fully winged species from southeast Asia, and Sia (Maxentius) with four wingless species from southern Africa. Because there is a dearth of published data about the behavior and biology of these insects, we present new field and laboratory research on southern African Sia (Maxentius), gather museum and literature information, and present guidelines for collecting and rearing specimens. While we make no taxonomic decisions, this review should be useful for future studies, including a needed taxonomic revision. We also compare results from these southern African JCs with recent investigations on related New World taxa, where fascinating biological traits and extensive cryptic biodiversity have been uncovered. DNA analysis reveals that these Old and New World JCs are polyphyletic. Key words: Stenopelmatoidea, Siinae, South Africa, Maxentius Introduction As presently understood, the orthopteran superfamily Stenopelmatoidea (=Gryllacridoidea) consists of six families (Eades et al. -

Orthopteran Communities in the Conifer-Broadleaved Woodland Zone of the Russian Far East
Eur. J. Entomol. 105: 673–680, 2008 http://www.eje.cz/scripts/viewabstract.php?abstract=1384 ISSN 1210-5759 (print), 1802-8829 (online) Orthopteran communities in the conifer-broadleaved woodland zone of the Russian Far East THOMAS FARTMANN, MARTIN BEHRENS and HOLGER LORITZ* University of Münster, Institute of Landscape Ecology, Department of Community Ecology, Robert-Koch-Str. 26, D-48149 Münster, Germany; e-mail: [email protected] Key words. Orthoptera, cricket, grasshopper, community ecology, disturbance, grassland, woodland zone, Lazovsky Reserve, Russian Far East, habitat heterogeneity, habitat specifity, Palaearctic Abstract. We investigate orthopteran communities in the natural landscape of the Russian Far East and compare the habitat require- ments of the species with those of the same or closely related species found in the largely agricultural landscape of central Europe. The study area is the 1,200 km2 Lazovsky State Nature Reserve (Primorsky region, southern Russian Far East) 200 km east of Vladi- vostok in the southern spurs of the Sikhote-Alin Mountains (134°E/43°N). The abundance of Orthoptera was recorded in August and September 2001 based on the number present in 20 randomly placed 1 m² quadrates per site. For each plot (i) the number of species of Orthoptera, (ii) absolute species abundance and (iii) fifteen environmental parameters characterising habitat structure and micro- climate were recorded. Canonical correspondence analysis (CCA) was used first to determine whether the Orthoptera occur in ecol- ogically coherent groups, and second, to assess their association with habitat characteristics. In addition, the number of species and individuals in natural and semi-natural habitats were compared using a t test. -

Phylogeny of Ensifera (Hexapoda: Orthoptera) Using Three Ribosomal Loci, with Implications for the Evolution of Acoustic Communication
Molecular Phylogenetics and Evolution 38 (2006) 510–530 www.elsevier.com/locate/ympev Phylogeny of Ensifera (Hexapoda: Orthoptera) using three ribosomal loci, with implications for the evolution of acoustic communication M.C. Jost a,*, K.L. Shaw b a Department of Organismic and Evolutionary Biology, Harvard University, USA b Department of Biology, University of Maryland, College Park, MD, USA Received 9 May 2005; revised 27 September 2005; accepted 4 October 2005 Available online 16 November 2005 Abstract Representatives of the Orthopteran suborder Ensifera (crickets, katydids, and related insects) are well known for acoustic signals pro- duced in the contexts of courtship and mate recognition. We present a phylogenetic estimate of Ensifera for a sample of 51 taxonomically diverse exemplars, using sequences from 18S, 28S, and 16S rRNA. The results support a monophyletic Ensifera, monophyly of most ensiferan families, and the superfamily Gryllacridoidea which would include Stenopelmatidae, Anostostomatidae, Gryllacrididae, and Lezina. Schizodactylidae was recovered as the sister lineage to Grylloidea, and both Rhaphidophoridae and Tettigoniidae were found to be more closely related to Grylloidea than has been suggested by prior studies. The ambidextrously stridulating haglid Cyphoderris was found to be basal (or sister) to a clade that contains both Grylloidea and Tettigoniidae. Tree comparison tests with the concatenated molecular data found our phylogeny to be significantly better at explaining our data than three recent phylogenetic hypotheses based on morphological characters. A high degree of conflict exists between the molecular and morphological data, possibly indicating that much homoplasy is present in Ensifera, particularly in acoustic structures. In contrast to prior evolutionary hypotheses based on most parsi- monious ancestral state reconstructions, we propose that tegminal stridulation and tibial tympana are ancestral to Ensifera and were lost multiple times, especially within the Gryllidae. -

Odonata: Aeshnidae): a Study of Traits from Larval Development to Adults
EUROPEAN JOURNAL OF ENTOMOLOGYENTOMOLOGY ISSN (online): 1802-8829 Eur. J. Entomol. 116: 269–280, 2019 http://www.eje.cz doi: 10.14411/eje.2019.031 ORIGINAL ARTICLE Biometry of the large dragonfl y Anax imperator (Odonata: Aeshnidae): A study of traits from larval development to adults MARCEAU MINOT 1, MICKAËL LE GALL2 and AURÉLIE HUSTÉ 1 1 Université de Rouen - ECODIV, Bat Blondel, Place Emile Blondel, Mont-Saint-Aignan 76821, France; e-mails: [email protected], [email protected] 2 IRSTEA, UR RIVERLY, Centre de Lyon – Villeurbanne, 69625 Villeurbanne Cedex, France; e-mail: [email protected] Key words. Odonata, Aeshnidae, Anax imperator, body length, body mass, larval rearing, sexual size dimorphism, traits Abstract. Insect larval development affects adult traits but the biometric relationships are usually poorly understood, including large odonates. In this study, measurements of morphological traits of larvae, exuviae and adults of Anax imperator were recorded. They were used to investigate the effects of early development on adult morphology. Results showed an increase in larval length during the fi nal instar and the length of its exuviae signifi cantly exceeded that of the larva. Length and body mass of teneral adults were strongly related to the length of their exuviae. Adult males were signifi cantly longer than adult females, while both had the same body mass at emergence. Length of teneral adults was negatively related to the date of emergence in both sexes. During maturation, body mass of males only increased slightly whereas that of females increased greatly. Mature specimens were also signifi cantly longer than teneral individuals. -

Barcode of Life Data Systems (BOLD) Versus Genbank Molecular Identification of a Dragonfly from the UAE in Comparison to the Morphological Identification
OnLine Journal of Biological Sciences Original Research Paper Barcode of Life Data Systems (BOLD) Versus GenBank Molecular Identification of a Dragonfly from the UAE in Comparison to the Morphological Identification 1Noora Almansoori, 1,2Mohamed Rizk Enan and 1Mohammad Ali Al-Deeb 1Department of Biology, United Arab Emirates University, Al-Ain, UAE 2Agricultural Research Center, Agricultural Genetic Engineering Research Institute, Giza, Egypt Article history Abstract: Dragonflies are insects in the order Odonata. They inhabit Received: 26-09-2019 freshwater ecosystems and are found in the UAE. To date, few checklists Revised: 19-11-2019 have been published for the local dragonflies and the used identification Accepted: 29-11-2019 keys are not comprehensive of Arabia. The aim of this study was to provide a molecular identification of a dragonfly based on the mitochondrial Corresponding Author: Cytochrome c Oxidase subunit I (COI) gene using the National Center for Mohammad Ali Al-Deeb, Biotechnology Information (NCBI) database and the Barcode of Life Data Department of Biology, United Systems (BOLD) in comparison with the morphology. The insect’s DNA Arab Emirates University, Al- was extracted and the PCR was performed on the target gene. The insect Ain, UAE Email: [email protected] was identified initially as Anax imperator based on the NCBI database and as Anax parthenope based on the BOLD. However, the morphological identification was in agreement with the one produced by the BOLD. The results of this study is a demonstration of how, in some cases, the DNA- based identification does not provide a conclusive species designation and that a morphology-based identification is needed. -

Aliens: the Invasive Species Bulletin Newsletter of the IUCN/SSC Invasive Species Specialist Group
Aliens: The Invasive Species Bulletin Newsletter of the IUCN/SSC Invasive Species Specialist Group ISSN 1173-5988 Issue Number 31, 2011 Coordinator CONTENTS Piero Genovesi, ISSG Chair, ISPRA Editors Editorial pg. 1 Piero Genovesi and Riccardo Scalera News from the ISSG pg. 2 Assistant Editor ...And other news pg. 4 Anna Alonzi Monitoring and control modalities of a honeybee predator, the Yellow Front Cover Photo legged hornet Vespa velutina The yellow-legged hornet Vespa velutina nigrithorax (Hymenoptera: © Photo by Quentin Rome Vespidae) pg. 7 Improving ant eradications: details of more successes, The following people a global synthesis contributed to this issue and recommendations pg. 16 Shyama Pagad, Carola Warner Introduced reindeer on South Georgia – their impact and management pg. 24 Invasive plant species The newsletter is produced twice a year and in Asian elephant habitats pg. 30 is available in English. To be added to the AlterIAS: a LIFE+ project to curb mailing list, or to download the electronic the introduction of invasive version, visit: ornamental plants in Belgium pg. 36 www.issg.org/newsletter.html#Aliens Investigation of Invasive plant Please direct all submissions and other ed- species in the Caucasus: itorial correspondence to Riccardo Scalera current situation pg. 42 [email protected] The annual cost of invasive species to the British economy quantified pg. 47 Published by Eradication of the non-native ISPRA - Rome, Italy sea squirt Didemnum vexillum Graphics design from Holyhead Harbour, Wales, UK pg. 52 Franco Iozzoli, ISPRA Challenges, needs and future steps Coordination for managing invasive alien species Daria Mazzella, ISPRA - Publishing Section in the Western Balkan Region pg. -

© 2016 David Paul Moskowitz ALL RIGHTS RESERVED
© 2016 David Paul Moskowitz ALL RIGHTS RESERVED THE LIFE HISTORY, BEHAVIOR AND CONSERVATION OF THE TIGER SPIKETAIL DRAGONFLY (CORDULEGASTER ERRONEA HAGEN) IN NEW JERSEY By DAVID P. MOSKOWITZ A dissertation submitted to the Graduate School-New Brunswick Rutgers, The State University of New Jersey In partial fulfillment of the requirements For the degree of Doctor of Philosophy Graduate Program in Entomology Written under the direction of Dr. Michael L. May And approved by _____________________________________ _____________________________________ _____________________________________ _____________________________________ New Brunswick, New Jersey January, 2016 ABSTRACT OF THE DISSERTATION THE LIFE HISTORY, BEHAVIOR AND CONSERVATION OF THE TIGER SPIKETAIL DRAGONFLY (CORDULEGASTER ERRONEA HAGEN) IN NEW JERSEY by DAVID PAUL MOSKOWITZ Dissertation Director: Dr. Michael L. May This dissertation explores the life history and behavior of the Tiger Spiketail dragonfly (Cordulegaster erronea Hagen) and provides recommendations for the conservation of the species. Like most species in the genus Cordulegaster and the family Cordulegastridae, the Tiger Spiketail is geographically restricted, patchily distributed with its range, and a habitat specialist in habitats susceptible to disturbance. Most Cordulegastridae species are also of conservation concern and the Tiger Spiketail is no exception. However, many aspects of the life history of the Tiger Spiketail and many other Cordulegastridae are poorly understood, complicating conservation strategies. In this dissertation, I report the results of my research on the Tiger Spiketail in New Jersey. The research to investigate life history and behavior included: larval and exuvial sampling; radio- telemetry studies; marking-resighting studies; habitat analyses; observations of ovipositing females and patrolling males, and the presentation of models and insects to patrolling males. -

Discovery of an Isolated Population of Anax Longipes in Michigan (Odonata: Aeshnidae)
The Great Lakes Entomologist Volume 29 Number 3 - Fall 1996 Number 3 - Fall 1996 Article 6 October 1996 Discovery of an Isolated Population of Anax Longipes in Michigan (Odonata: Aeshnidae) Michael A. Kielb University of Michigan Mark F. O'Brien University of Michigan Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Kielb, Michael A. and O'Brien, Mark F. 1996. "Discovery of an Isolated Population of Anax Longipes in Michigan (Odonata: Aeshnidae)," The Great Lakes Entomologist, vol 29 (3) Available at: https://scholar.valpo.edu/tgle/vol29/iss3/6 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Kielb and O'Brien: Discovery of an Isolated Population of <i>Anax Longipes</i> in Mi 1996 THE GREAT LAKES ENTOMOLOGIST 161 DISCOVERY OF AN ISOLATED POPULATION OF ANAX LONGIPES IN MICHIGAN (ODONATA: AESHNIDAE) Michael A. Kielb and Mark F. O'Brien 1 ABSTRACT Anax longipes is a large aeshnid dragonfly previously unknown from Michigan. Adults and larvae were found in abundance at a series of experi mental ponds within the E.S. George Reserve in Livingston County, Michi gan. Anax longipes Hagen was previously undocumented from Michigan (Kor mondy 1958), and is rarely encountered in the Great Lakes Region (Glotzhober 1995, Kielb 1996, Tennessen 1992). The species is widely distrib uted throughout the eastern United States, from Massachusetts to Wisconsin and south to Florida, with scattered records from southern Ontario, Canada (Dunkle 1989). -

Dragonflies at All, Though
Odonatologica 5 (I): 1■ 10 March I, 1976 PROCEEDINGS OF THE THIRD INTERNATIONAL SYMPOSIUM OF ODONATOLOGY Lancaster, July 14-18, 1975 Part I A history of odonatologyin the British Isles* R.M. Gambles Windings, Whitchurch Hill, Reading RG8 7NU, England Received and Accepted December 1, 1975 The development of odonatology in the British Isles is traced from 1634 to the present and reference is made to the contributions made by British odonatologists to the knowledge of the fauna of other regions and to odonato- in the the those of K.J. logy general. Among portraits accompanying paper, MORTON (1858-1940), H. CAMPION (1869-1932), and N. MacNEILL (1899-1969)had not been published previously. I have been asked to give a brief account of the history of odonatology in the who British Isles and of British odonatologists. There are a number here present are eminently more qualified than I am for this task. But doubtless modesty would prevent them from doing justice to their own contributionto the Science. 1 Whereas I myself, having contributed nothing, need have no such inhibitions. Probably the earliest publication in this country to dealwith dragonflies at all, though they formed only a very small part of the whole„was ’Tnsectorum Theatrum” 1634, by Dr THOMAS MOUFFET (1533-1604). He did not name but them into any dragonflies, attempted a very rough classification, dividing maxima, media and minima, roughly corresponding with the familiesAeshnidae, , and the and his Libellulidae, Zygoptera, among primitive figures we can recog- nise Libellula and It contribution to depressa Calopteryx virgo. was not a great but least our Science, it was at a beginning. -
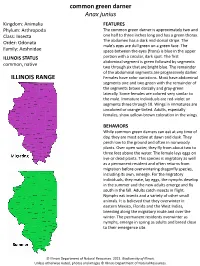
Common Green Darner Anax Junius ILLINOIS RANGE
common green darner Anax junius Kingdom: Animalia FEATURES Phylum: Arthropoda The common green darner is approximately two and Class: Insecta one half to three inches long and has a green thorax. Order: Odonata The abdomen has a dark mid-dorsal stripe. The male’s eyes are dull green on a green face. The Family: Aeshnidae space between the eyes (frons) is blue in the upper ILLINOIS STATUS portion with a circular, dark spot. The first abdominal segment is green followed by segments common, native two through six that are bright blue. The remainder of the abdominal segments are progressively darker. ILLINOIS RANGE Females have color variations. Most have abdominal segments one and two green with the remainder of the segments brown dorsally and gray-green laterally. Some females are colored very similar to the male. Immature individuals are red-violet on segments three through 10. Wings in immatures are uncolored or orange-tinted. Adults, especially females, show yellow-brown coloration in the wings. BEHAVIORS While common green darners can eat at any time of day, they are most active at dawn and dusk. They perch low to the ground and often in nonwoody plants. Over open water, they fly from about two to three feet above the water. The female lays eggs on live or dead plants. This species is migratory as well as a permanent resident and often returns from migration before overwintering dragonfly species, including its own, emerge. For the migratory individuals, they mate, lay eggs, the nymphs develop in the summer and the new adults emerge and fly south in the fall. -

International Revîëw of Poultry Science
TOME IX. 1936. No. 1/2 INTERNATIONAL REVÎËW OF POULTRY SCIENCE OFFICIAL ORGAN OF THE WORLD'S POULTRY SCIENCE ASSOCIATION ¡f73S EDITOR: Dr. B. J. C. TE HENNEPE ROTTERDAM (Holland) This Review is free fo all members of the World's Poultry Science Association. AH subscriptions should be sent to the Editor: Dr. B. J. C. te Hennepe, Rotterdam, or to the Secretary- Treasurer: Dr. G. F. Heuser, Cornell University, Ithaca, N.y., U.S.A. SUBSCRIPTIONS. $5.00 annually in advance. The personal membership of the W.P.S.A. per amounts to $5.00 year For affiliated societies „ „ $25.00 ADVERTISEMENT RATES. One page, per issue $12.00 Half page, per issue $7.00 NATIONAL POULTRY COUNCIL LAYING TRIALS REGISTER Vol. IX Containing Official Records obtained at RECOGNISED LAYING TRIALS Now on sale: Price 6d., postage paid. Copies of Vols. I-VIII are also on sale, price 6d. each, postage paid. Apply to: THE SECRETARY, National Poultry Council Avenue Chambers, 4, Vernon Place, London W. C. 1 ENGLAND Sir Edward Brown Above: Reidisminisfer for Agriculture and ßeidisbauernführer R. Walther Darré, President of the VI fh World's Poultry Congress. Left: Mr. Karl Vetter, Inspector General of the „Reichsnährstand" and President of the National Union of German Small Stock Breeders, Acting President of the VI th World's Poultry Congress. Below: Dr.Walther Kupsdi, ¡Secretary General of theVIfh World's Poultry Congress. Äf the reception of the Press in connection with the VI th World's Poultry Congress. The managing President of the Congress, Karl Vetter, explains to the Honorary President of the W.P.S.A., Sir Edward Brown, as well as to Ministerialdirektor Dr.