Diptera: Mycetophilidae) by William G
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
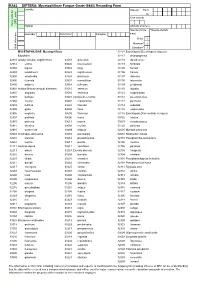
Ra82 Diptera
RA82 DIPTERA: Mycetophilinae Fungus Gnats (6480) Recording Form Locality Date(s) from: to: Vice county GPS users Grey cells for Habitat Altitude (metres) Source (circle *Source details Recorder Determiner Compiler one) Field 1 Museum* 2 Grid reference Literature* 3 MYCETOPHILIDAE: Mycetophilinae 33101 Exechiopsis (Exechiopsis) clypeata Exechiini 33117 dryaspagensis 32801 Allodia (Allodia) anglofennica 33519 griseolum33118 dumitrescae 32912 embla 33526 intermedium33119 fimbriata 32802 lugens 33522 kingi33120 furcata 32803 lundstroemi 33523 nigrofuscum33106 hammi 32804 ornaticollis 33524 proximum33107 indecisa 32806 truncata 33527 rosmellitum33108 intersecta 32805 zaitzevi 33514 ruficorne33109 jenkinsoni 32901 Allodia (Brachycampta) alternans 33515 serenum33110 ligulata 32910 angulata 33516 sericoma33121 magnicauda 32903 barbata 33601 Cordyla brevicornis33112 pseudindecisa 32904 czernyi 33602 crassicornis33113 pulchella 32909 foliifera 33603 fasciata33114 subulata 32905 grata 33604 fissa33115 unguiculata 32906 neglecta 33605 flaviceps33116 Exechiopsis (Xenexechia) crucigera 32907 pistillata 33606 fusca33002 leptura 32915 protenta 33613 insons33003 membranacea 32914 silvatica 33608 murina33122 pollicata 32916 westerholti 33609 nitidula32605 Myrosia maculosa 32602 Allodiopsis domestica 33610 parvipalpis32601 Notolopha cristata 32610 korolevi 33614 pseudomurina32701 Pseudexechia aurivernica 32607 rustica 33611 pusilla32706 monica 32212 Anatella alpina 33612 semiflava32705 parallela 32213 ankeli 33201 Exechia bicincta32703 trisignata 32216 bremia -

Impact of Imidacloprid and Horticultural Oil on Nonâ•Fitarget
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Masters Theses Graduate School 8-2007 Impact of Imidacloprid and Horticultural Oil on Non–target Phytophagous and Transient Canopy Insects Associated with Eastern Hemlock, Tsuga canadensis (L.) Carrieré, in the Southern Appalachians Carla Irene Dilling University of Tennessee - Knoxville Follow this and additional works at: https://trace.tennessee.edu/utk_gradthes Part of the Entomology Commons Recommended Citation Dilling, Carla Irene, "Impact of Imidacloprid and Horticultural Oil on Non–target Phytophagous and Transient Canopy Insects Associated with Eastern Hemlock, Tsuga canadensis (L.) Carrieré, in the Southern Appalachians. " Master's Thesis, University of Tennessee, 2007. https://trace.tennessee.edu/utk_gradthes/120 This Thesis is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Masters Theses by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a thesis written by Carla Irene Dilling entitled "Impact of Imidacloprid and Horticultural Oil on Non–target Phytophagous and Transient Canopy Insects Associated with Eastern Hemlock, Tsuga canadensis (L.) Carrieré, in the Southern Appalachians." I have examined the final electronic copy of this thesis for form and content and recommend that it be accepted in partial fulfillment of the equirr ements for the degree of Master of Science, with a major in Entomology and Plant Pathology. Paris L. Lambdin, Major Professor We have read this thesis and recommend its acceptance: Jerome Grant, Nathan Sanders, James Rhea, Nicole Labbé Accepted for the Council: Carolyn R. -

Recent Noteworthy Findings of Fungus Gnats from Finland and Northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae)
Biodiversity Data Journal 2: e1068 doi: 10.3897/BDJ.2.e1068 Taxonomic paper Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae) Jevgeni Jakovlev†, Jukka Salmela ‡,§, Alexei Polevoi|, Jouni Penttinen ¶, Noora-Annukka Vartija# † Finnish Environment Insitutute, Helsinki, Finland ‡ Metsähallitus (Natural Heritage Services), Rovaniemi, Finland § Zoological Museum, University of Turku, Turku, Finland | Forest Research Institute KarRC RAS, Petrozavodsk, Russia ¶ Metsähallitus (Natural Heritage Services), Jyväskylä, Finland # Toivakka, Myllyntie, Finland Corresponding author: Jukka Salmela ([email protected]) Academic editor: Vladimir Blagoderov Received: 10 Feb 2014 | Accepted: 01 Apr 2014 | Published: 02 Apr 2014 Citation: Jakovlev J, Salmela J, Polevoi A, Penttinen J, Vartija N (2014) Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae). Biodiversity Data Journal 2: e1068. doi: 10.3897/BDJ.2.e1068 Abstract New faunistic data on fungus gnats (Diptera: Sciaroidea excluding Sciaridae) from Finland and NW Russia (Karelia and Murmansk Region) are presented. A total of 64 and 34 species are reported for the first time form Finland and Russian Karelia, respectively. Nine of the species are also new for the European fauna: Mycomya shewelli Väisänen, 1984,M. thula Väisänen, 1984, Acnemia trifida Zaitzev, 1982, Coelosia gracilis Johannsen, 1912, Orfelia krivosheinae Zaitzev, 1994, Mycetophila biformis Maximova, 2002, M. monstera Maximova, 2002, M. uschaica Subbotina & Maximova, 2011 and Trichonta palustris Maximova, 2002. Keywords Sciaroidea, Fennoscandia, faunistics © Jakovlev J et al. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. -

Of Greece, Its Islands
CHANDLERet al.: 255-314 - Studia dipterologica 12 (2005) Heft 2 ISSN 0945-3954 The Fungus Gnats (Diptera: Bolitophilidae, Diadocidiidae, Ditomyiidae , Keroplatidae and Mycetophilidae) of Greece, its islands and Cyprus [Die Pilzmiicken (Diptera: Bolitophilidae, Diadocidiidae, Ditomyiidae, Keroplatidae und Mycetophilidae) Griechenlands und seiner Inseln sowie Zypern4 1 by Peter J. CHANDLER, Dimitar N. BECHEV and Norbert CASPERS Mclksham (UK) Plovdiv (Bulgaria) Bechen (Gernlany) - - -. - ~ Abstract The spccics of fungu\ gnats (Bolitophilidae, Diadoc~dildae,Ditomyiidac. Keroplat~d:~eand Mycetophilidae) o~urringin Greece and Cyprus are reviewed. Altogether 201 species :Ire recorded, 189 for Greece and 69 for Cyprus. Of these 126 specie5 arc newly recorded fol. Greece and 36 arc newly recorded for Cyprus. The following new taxa arc described from Greece: Macrorrhyrtcha ibis spec. nov., M. pelargos spec. nov., M. laconica spec. nov., Macrocera critica spec. nov., Docosia cephaloniae spec. nov., D. enos spec. nov., D. pa- siphae spec. nov., Megophthalmidia illyrica spec. nov.. M. ionica spec. nov., M. pytho spec. nov., Mycomya thrakis spec. nov., Allocolocera scheria spec. nov., Sciophila pandora spec. nov., Ryrnosia labyrinthos spec. nov.; M. ill\,ric,cr is also recorded troln Croc~lia.The follow- ing ncw taa are described from Cyprus: Macrocera cypriaca spec. nov., Megophthalmidia alrzicola spec. nov., M. cedricola spec. nov. The following neu synonymies are propod: M!,c,c~r~iwrenuis I WXLKER,1856) = M. interniissa PL.ASSMA~N,l984 syn. nov., Plrror~rtr~1.illi.s- torri DLIFI>ZICKI,1889 = P rnciscr CASFERS,1991 syn. nov. A key is provided for thc western Palaearctic specie5 of M(ic-i.orrh~~~ic-IrciWI~~ERTZ. -

Zootaxa, the Fungus Gnats (Diptera: Bolitophilidae
Zootaxa 2318: 450–506 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) The fungus gnats (Diptera: Bolitophilidae, Keroplatidae, Mycetophilidae) of Sardinia, with description of six new species* PETER J. CHANDLER 606B Berryfield Lane, Melksham, Wilts SN12 6EL, United Kingdom. E-mail: [email protected] *In: Cerretti, P., Mason, F., Minelli, A., Nardi, G. & Whitmore, D. (Eds), Research on the Terrestrial Arthropods of Sardinia (Italy). Zootaxa, 2318, 1–602. Table of contents Abstract . .450 Introduction . .451 Study area . .452 Material and methods . .452 Abbreviations . .453 Sampling sites . .454 Faunistic list . .456 Bolitophilidae . .456 Keroplatidae, Keroplatinae, Keroplatini . .456 Orfeliini . .457 Macrocerinae . .462 Mycetophilidae, Gnoristinae . .465 Leiinae . .469 Mycetophilinae, Exechiini . .472 Mycetophilini . .480 Mycomyinae . .492 Sciophilinae . .495 Discussion . .500 Acknowledgements . .501 References . .502 Abstract The fungus gnat fauna of Sardinia is reviewed and data presented for all species recorded. Altogether one species of Bolitophilidae, 16 species of Keroplatidae and 105 species of Mycetophilidae are recognised as occurring in Sardinia. As the bolitophilid and two of the mycetophilid species are represented only by females and are not determined to species level, the total confirmed Sardinian list stands at 119 species. Four species of Keroplatidae and 19 species of Mycetophilidae are new to the total Italian fauna, whereas three species of Keroplatidae and 32 species of Mycetophilidae are newly recorded for the island of Sardinia. Six species are described as new to science: two Keroplatidae (Urytalpa juliae sp. nov., Macrocera nuragica sp. nov.) and four Mycetophilidae (Boletina ichnusa sp. nov., Trichonta sandalyon sp. -

Zootaxa 3549: 1–117 (2012) ISSN 1175-5326 (Print Edition) ZOOTAXA Copyright © 2012 · Magnolia Press Monograph ISSN 1175-5334 (Online Edition)
Zootaxa 3549: 1–117 (2012) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA Copyright © 2012 · Magnolia Press Monograph ISSN 1175-5334 (online edition) urn:lsid:zoobank.org:pub:2412CB4F-4D29-4988-80C1-205D16767678 ZOOTAXA 3549 Systematics and Phylogeny of Leptomorphus Curtis (Diptera: Mycetophilidae) CHRISTOPHER J. BORKENT1 & TERRY A. WHEELER2 1 Plant Pest Diagnostics Center, California Department of Food and Agriculture, 3294 Meadowview Road, Sacramento, CA 95832, USA. Corresponding author, email: [email protected] 2Dept. Natural Resource Sciences, McGill University, Macdonald Campus, Ste-Anne-de-Bellevue, QC H9X 3V9 CANADA. email: [email protected] Magnolia Press Auckland, New Zealand Accepted by P.H. Kerr: 6 Sept. 2012; published: 15 Nov. 2012 CHRISTOPHER J. BORKENT & TERRY A. WHEELER Systematics and Phylogeny of Leptomorphus Curtis (Diptera: Mycetophilidae) (Zootaxa 3549) 117 pp.; 30 cm. 15 Nov 2012 ISBN 978-1-77557-014-1 (paperback) ISBN 978-1-77557-015-8 (Online edition) FIRST PUBLISHED IN 2012 BY Magnolia Press P.O. Box 41-383 Auckland 1346 New Zealand e-mail: [email protected] http://www.mapress.com/zootaxa/ © 2012 Magnolia Press All rights reserved. No part of this publication may be reproduced, stored, transmitted or disseminated, in any form, or by any means, without prior written permission from the publisher, to whom all requests to reproduce copyright material should be directed in writing. This authorization does not extend to any other kind of copying, by any means, in any form, and for any purpose other than private research use. ISSN 1175-5326 (Print edition) ISSN 1175-5334 (Online edition) 2 · Zootaxa 3549 © 2012 Magnolia Press BORKENT & WHEELER Table of contents Abstract . -

Awenda Provincial Park
AWENDA PROVINCIAL PARK One Malaise trap was deployed at Awenda Provincial Park in 2014 (44.82534, -79.98458, 231m ASL; Figure 1). This trap collected arthropods for twenty weeks from April 29 – September 19, 2014. All 10 Malaise trap samples were processed; every other sample was analyzed using the individual specimen protocol while the second half was analyzed via bulk analysis. A total of 3029 BINs were obtained. Over half the BINs captured were flies (Diptera), followed by bees, ants and wasps (Hymenoptera), moths and butterflies (Lepidoptera), and true bugs (Hemiptera; Figure 2). In total, 595 arthropod species were named, representing 21.3% of the BINs from the Figure 1. Malaise trap deployed at Awenda Provincial site (Appendix 1). All the BINs were assigned at least Park in 2014. to family, and 54% were assigned to a genus (Appendix 2). Specimens collected from Awenda represent 214 different families and 705 genera. Diptera Hymenoptera Lepidoptera Hemiptera Coleoptera Trombidiformes Sarcoptiformes Psocodea Mesostigmata Araneae Entomobryomorpha Mecoptera Symphypleona Trichoptera Neuroptera Thysanoptera Dermaptera Pseudoscorpiones Stylommatophora Odonata Opiliones Orthoptera Figure 2. Taxonomy breakdown of BINs captured in the Malaise trap at Awenda. APPENDIX 1. TAXONOMY REPORT Class Order Family Genus Species Arachnida Araneae Agelenidae Agelenopsis Clubionidae Clubiona Clubiona kastoni Dictynidae Emblyna Emblyna sublata Linyphiidae Ceraticelus Ceraticelus atriceps Ceraticelus fissiceps Ceratinella Ceratinella brunnea Ceratinops -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

Study of Systemic Status
http://www.paper.edu.cn 浙 江 林 学 院 学 报 2003,20(1):32—36 Journa l of Zh ejiang F orestry C ollege A rticle ID :1000—5692(2003)01—0032—05 S tud y of system ic statu s of M yCetO p h -Iida e W A N G Y i—ping ,W U H ong ,X U H ua—chao (Institute of Forest Protection ,Zhejiang Forestry C ollege ,L in’an 311300 .Zhejiang .C hina ) A bstract:T he actuality of M ycetoph ilidae fam ily research has been illustrated in detail. T11e research includes biology , classification an d idenfication , geograp hical distribution , status of system ic classification and system developm ent. Th e existed prob lem s and expectation are also explored in the pap er .A ccordin g to the latest inform ation , M ycetop hilidae fam ily is com p osed of 5 subfam ilies , i. e , M ycom yinae, Sciophilinae, G noristinae, Leiinae and M ycetophilinae . M ycetophilinae includes 2 tribes :E xechiini an d M ycetophilini. R efeFences 2 8 K ey w o rd s : M ycetophilidae ;geograp hical d istrib ution ; clas sification status CLC Number:Q969.44 Document Code:A Th e fami ly M ycetoph ilidae belon gs to the superfam ily S ciaroidea in the order D iptera , w h ich is th e largest fami ly in the superfami ly S ciaroidea⋯ . B ecause of this insects feeding on edible fungi or large fungus body , a lot of scholars have been involved in th e research on fungu s pest species , and great progress in research on fungu s gn ats has b een m ade . B ased on research results an d literature published at hom e and ab road in the pas t tw o decades , the authors m ak e a sum m arization on the -

Zootaxa, Diptera, Mycetophilidae
Zootaxa 856: 1–35 (2005) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA 856 Copyright © 2005 Magnolia Press ISSN 1175-5334 (online edition) A review of fungus gnats in the tribe Exechiini (Diptera, Myceto- philidae) from the J. W. Zetterstedt collection at the Museum of Zoology in Lund, Sweden JOSTEIN KJÆRANDSEN Museum of Zoology, Lund University, Helgonavägen 3, S-223 62 Lund, Sweden. ([email protected]) Table of contents Abstract . 1 Introduction . 2 Material and methods . 4 Result . 6 List of species . 12 Acknowledgements . 33 References . 33 Abstract The collections of fungus gnats by Johan Wilhelm Zetterstedt (1785–1874), lodged in the Museum of Zoology in Lund, Sweden, are examined for all species belonging in the tribe Exechiini Edwards. The majority of the material was collected in Fennoscandia, mainly in Sweden, in the first half of the 19th century. Altogether 37 species of the tribe Exechiini could be safely identified. Three additional species are strongly indicated to be present in the collections, but could not be identified with certainty, viz. Allodia (Brachycampta) alternans (Zetterstedt, 1838), Cordyla murina Winnertz, 1863 and Stigmatomeria crassicornis (Stannius, 1831). Some of Zetterstedt's types have been erroneously synonymized and misinterpreted in modern literature. Hence, illustra- tions of terminalia are presented for all recognizable Exechiini types described by Zetterstedt. In order to preserve nomenclatural stability a lectotype is selected for Brevicornu griseolum (Zetter- stedt, 1852) sensu auctore nec Edwards, and a neotype is selected for Allodia (Brachycampta) alternans (Zetterstedt, 1838). Two species names are reinstated, viz. Brevicornu canescens (Zetter- stedt, 1852) sp. -

List of Talk Presentations, Alphabetically by Senior Author, English and French Titles and Abstracts
List of Talk Presentations, Alphabetically by Senior Author, English and French Titles and Abstracts Abdel Ghany, A.1; Awadalla, S.S.2; Abdel-Baky, N.F.3; EL- Syrafi, H.A.4; Fields, P. G.5 Economic Entomology Department, Faculty of Agriculture, Mansoura University, Mansoura 35516, Egypt. (1,2,3,4); Cereal Research Centre, Agriculture & Agri-Food Canada, 195 Dafoe Road, Winnipeg, MB, R3T 2M9 (5). E-mail:[email protected] (?); [email protected] (5)co-author e-mails Susceptibility of Stegobium paniceum (Coleoptera: Anobiidae) of different life stages to high temperature. The drugstore beetle, Stegobium paniceum, is a pest of stored medicinal and aromatic plants. The heat tolerance of different stages from highest to lowest is young larvae, old larvae, eggs, adult and pupae. Heat treatments that control young larvae should control all insect stages. The LT90 for young larvae at 42, 45, 50, 55 and 60°C is 13, 10, 1.4, 0.06 and 0.05h respectively. La susceptibilité de différents stades de Stegobium paniceum (Coleoptera: Anobiidae) à différentes températures. Le stégobie des pharmacies, Stegobium paniceum, est un ravageur de plantes médicinales et aromatiques entreposées. La tolérance à la chaleur dans les différents stades, en ordre décroissant, s’illustre comme suit: jeunes larves, larves âgées, adultes et pupes. Les traitements thermiques qui contrôlent les jeunes larves devraient donc contrôler tous les stades d’insectes. La LT90 des jeunes larves à 42, 45, 50, 55 et 60°C est de 13, 10, 1.4, 0.06 et 0.05 respectivement. Abele, Suzanne1; Macdonald, Ellen2; Spence, John3 University of Alberta, Edmonton, AB T6G 2R3 (1,2,3). -

Holland Landing Prairie Provincial Park
HOLLAND LANDING PRAIRIE PROVINCIAL PARK One Malaise trap was deployed at Holland Landing Prairie Provincial Park in 2014 (44.11894, -79.48795, 271m ASL; Figure 1). This trap collected arthropods for twenty weeks from May 1 – September 22, 2014. All 10 Malaise trap samples were processed using the bulk sample analysis protocol. A total of 2345 BINs were obtained. Over half the BINs captured were flies (Diptera), followed by bees, ants and wasps (Hymenoptera), moths and butterflies (Lepidoptera), and true bugs (Hemiptera; Figure 2). In total, 544 arthropod species were named, representing 26.1% of the BINs from the site (Appendix 1). All the BINs were assigned at least to family, and 60.5% were assigned to a genus Figure 1. Malaise trap deployed at Holland Landing (Appendix 2). Specimens collected from Holland Prairie Provincial Park in 2014. Landing Prairie represent 165 different families and 623 genera. Figure 2. Taxonomy breakdown of BINs captured in the Malaise trap at Holland Landing Prairie. APPENDIX 1. TAXONOMY REPORT Class Order Family Genus Species Arachnida Araneae Theridiidae Theridion Theridion murarium Diplopoda Julida Julidae Insecta Psocodea Amphipsocidae Polypsocus Polypsocus corruptus Caeciliusidae Valenzuela Valenzuela burmeisteri Valenzuela flavidus Valenzuela nadleri Ectopsocidae Ectopsocus Ectopsocus meridionalis Lachesillidae Lachesilla Lepidopsocidae Echmepteryx Echmepteryx hageni Peripsocidae Peripsocus Peripsocus madidus Peripsocus subfasciatus Psocidae Cerastipsocus Trichadenotecnum Trichadenotecnum alexanderae