Genetic Structure and Gene Flows Within Horses: a Genealogical Study at the French Population Scale
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Multiple Choice Choose the Answer That Best Completes Each Statement Or Question
Name Date Hour 9 The Horse Industry Multiple Choice Choose the answer that best completes each statement or question. _______ 1. The horse was first domesticated in Europe and Asia about ____ . A. 1,000 years ago B. 3,000 years ago C. 5,000 years ago D. 8,000 years ago _______ 2. Horses were brought to the New World by Spanish explorers in the ____ . A. 15th century B. 16th century C. 17th century D. 18th century _______ 3. Horses are measured in terms of hands with a hand being ____ . A. two inches B. four inches C. six inches D. eight inches _______ 4. Ponies are shorter than horses and can be anywhere from 8 to ____ . A. 10 hands high B. 12.2 hands high C. 14.2 hands high D. 16 hands high _______ 5. Which horse breed is considered the oldest purebred horse in the world? A. Arabian B. Appaloosa C. Thoroughbred D. Quarter Horse Introduction to Agriscience | Unit 9 Test CIMC 1 _______ 6. Which horse breed has as one of its characteristics a distinctive spotted coat? A. Arabian B. Appaloosa C. Thoroughbred D. Quarter Horse _______ 7. Which horse breed was developed in the United States and got its name because of its great speed at short distances? A. Arabian B. Appaloosa C. Thoroughbred D. Quarter Horse _______ 8. Which horse breed was developed in the deserts of the Middle East? A. Arabian B. Appaloosa C. Thoroughbred D. Quarter Horse _______ 9. Which horse breed has a head characterized by a dished profile, prominent eye, large nostrils, and small muzzle? A. -

A Propos D'étalon
Association Nationale du Cheval Castillonnais d’Ariège Pyrénées Éviter la consanguinité et choisir le modèle de l’étalon Les goûts personnels et surtout ceux de la clientèle vont prévaloir : Il est cependant obligatoire de tenir compte du standard de la race. Au moment de choisir un étalon pour une jument déterminée, l’éleveur peut chercher soit à développer chez le produit à naître le maximum de qualités en additionnant celles du père et celles de la mère, soit à compenser les défauts de l’un(e) par les qualités de l’autre. Il convient également d’écarter tout accouplement qui ferait apparaître un ancêtre commun dans les 3 premières générations. Premier exemple : Cet accouplement est valable puisqu’il n’apparaît aucun ancêtre commun dans les 3 premières générations JOUR DE BREILH EMBRUN MARA Père NOBI CUMULUS DE GENTIANE BIGORRE MILHORAT VIOLETTE OLI DE HARDI CARBOUNEROS CIBADA Mère VOLUBILIS SILEX KALINKA TOSCA Deuxième exemple : Cet accouplement trop fortement consanguin doit être écarté. HAIDOUK DE QUEL POMPOM TOURNEMIRE ONEYL TEIGNE Père DU LOCH AR BIG JOUR DE BREILH BACALINE ? CUMULUS DE HARET DE BIGORRE BETHMALE CHIPIE Mère UNIQUE D’ALZEN HAIDOUK DE MARQUISE DES TOURNEMIRE VIELLES PRINCESSE 26/05/2011 1 / 5 Association Nationale du Cheval Castillonnais d’Ariège Pyrénées Au XXI ème siècle, l’informatique peut nous aider à éviter des accouplements consanguins. Voir : http://www.chevalcastillonnais.fr/public/simul.php A propos d’étalons Castillonnais Grâce aux pointages, nous avons la chance de disposer aujourd’hui d’une indexation des reproducteurs mâles. Pour chacun des étalons et chacun des postes morphologiques on sait aujourd’hui qui améliore ou détériore quoi. -

Genome-Wide Analysis Reveals Selection for Important Traits In
Genome-wide analysis reveals selection for important traits in domestic horse breeds Jessica L Petersen, James R Mickelson, Aaron K Rendahl, Stephanie J Valberg, Lisa S Andersson, Jeanette Axelsson, Ernie Bailey, Danika Bannasch, Matthew M Binns, Alexandre S Borges, et al. To cite this version: Jessica L Petersen, James R Mickelson, Aaron K Rendahl, Stephanie J Valberg, Lisa S Andersson, et al.. Genome-wide analysis reveals selection for important traits in domestic horse breeds. PLoS Genetics, Public Library of Science, 2013, 9, online (1), Non paginé. 10.1371/journal.pgen.1003211. hal-01000690 HAL Id: hal-01000690 https://hal.archives-ouvertes.fr/hal-01000690 Submitted on 29 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Genome-Wide Analysis Reveals Selection for Important Traits in Domestic Horse Breeds Jessica L. Petersen1*, James R. Mickelson1, Aaron K. Rendahl2, Stephanie J. Valberg1, Lisa S. Andersson3, Jeanette Axelsson3, Ernie Bailey4, Danika Bannasch5, Matthew M. Binns6, Alexandre S. Borges7, Pieter Brama8, Artur da Caˆmara Machado9, Stefano Capomaccio10, Katia Cappelli10, E. Gus Cothran11, Ottmar Distl12, Laura Fox-Clipsham13, Kathryn T. Graves4,Ge´rard Gue´rin14, Bianca Haase15, Telhisa Hasegawa16, Karin Hemmann17, Emmeline W. -
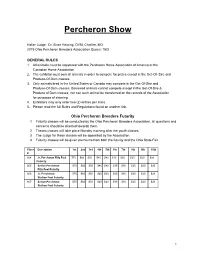
Percheron Show
Percheron Show Halter Judge: Dr. Brian Heuring, DVM, Chaffee, MO 2019 Ohio Percheron Breeders Association Queen: TBD GENERAL RULES 1. All animals must be registered with the Percheron Horse Association of America or the Canadian Horse Association 2. The exhibitor must own all animals in order to compete for prizes except in the Get-Of- Sire and Produce-Of-Dam classes. 3. Only animals bred in the United States or Canada may compete in the Get-Of-Sire and Produce-Of-Dam classes. Borrowed animals cannot compete except in the Get-Of-Sire & Produce of Dam classes, nor can such animal be transferred on the records of the Association for purposes of showing. 4. Exhibitors may only enter two (2) entries per class. 5. Please read the full Rules and Regulations found on another link. Ohio Percheron Breeders Futurity 1. Futurity classes will be conducted by the Ohio Percheron Breeders Association, all questions and concerns should be directed towards them. 2. Theses classes will take place Monday morning after the youth classes. 3. The Judge for these classes will be appointed by the Association. 4. Futurity classes will be given premiums from both the futurity and the Ohio State Fair Class Description 1st 2nd 3rd 4th 5th 6th 7th 8th 9th 10th # 604 Jr. Percheron Filly Foal $75 $60 $50 $45 $40 $35 $30 $25 $20 $20 Futurity 605 Senior Percheron $75 $60 $50 $45 $40 $35 $30 $25 $20 $20 Filly Foal Futurity 606 Jr. Percheron $75 $60 $50 $45 $40 $35 $30 $25 $20 $20 Stallion Foal Futurity 607 Senior Percheron $75 $60 $50 $45 $40 $35 $30 $25 $20 $20 Stallion Foal Futurity 1 PERCHERON HALTER CLASSES STALLIONS Entry Class Prize Description 1st 2nd 3rd 4th 5th 6th 7th 8th 9th 10th # Money Fee 625 $240 Registered Percheron Stallion, 3 Years $5 $75 $60 $50 $35 $20 & Over. -

List of Horse Breeds 1 List of Horse Breeds
List of horse breeds 1 List of horse breeds This page is a list of horse and pony breeds, and also includes terms used to describe types of horse that are not breeds but are commonly mistaken for breeds. While there is no scientifically accepted definition of the term "breed,"[1] a breed is defined generally as having distinct true-breeding characteristics over a number of generations; its members may be called "purebred". In most cases, bloodlines of horse breeds are recorded with a breed registry. However, in horses, the concept is somewhat flexible, as open stud books are created for developing horse breeds that are not yet fully true-breeding. Registries also are considered the authority as to whether a given breed is listed as Light or saddle horse breeds a "horse" or a "pony". There are also a number of "color breed", sport horse, and gaited horse registries for horses with various phenotypes or other traits, which admit any animal fitting a given set of physical characteristics, even if there is little or no evidence of the trait being a true-breeding characteristic. Other recording entities or specialty organizations may recognize horses from multiple breeds, thus, for the purposes of this article, such animals are classified as a "type" rather than a "breed". The breeds and types listed here are those that already have a Wikipedia article. For a more extensive list, see the List of all horse breeds in DAD-IS. Heavy or draft horse breeds For additional information, see horse breed, horse breeding and the individual articles listed below. -

WILL HE >JUSTIFY= the HYPE?
SATURDAY, APRIL 7, 2018 BLUE GRASS COULD BE WHITE OUT WILL HE >JUSTIFY= Assuming a potential snowstorm Friday night doesn=t put a THE HYPE? damper on things, a full field is set to go postward in Saturday=s GII Toyota Blue Grass S. The conversation about contenders must start with $1-million KEESEP yearling Good Magic (Curlin), who broke his maiden emphatically in the GI Breeders= Cup Juvenile at Del Mar en route to Eclipse Award honors. Heavily favored in the GII Xpressbet Fountain of Youth on seasonal debut at Gulfstream Mar. 3, he settled for a non-threatening third with no obvious excuse. Flameaway (Scat Daddy), second in the Mar. 10 GII Tampa Bay Derby behind expected scratch Quip (Distorted Humor) after annexing the GIII Sam F. Davis S. there in February, took a rained-off renewal of the GIII Bourbon S. in the Keeneland slop last October. Cont. p3 Justify | Benoit Photo IN TDN EUROPE TODAY ASTUTE AGENT HAS GLOBAL PERSPECTIVE >TDN Rising Star= Justify (Scat Daddy), almost certainly the Kelsey Riley sat down with Australia based Frenchman most highly regarded horse in the world to have not yet taken Louis Le Metayer to get his thoughts on various aspects of on stakes company, will get his class test on Saturday when he the Australian racing scene. Click or tap here to go straight faces the likes of MGISW Bolt d=Oro (Medaglia d=Oro) in the to TDN Europe. GI Santa Anita Derby. A 9 1/2-length jaw-dropping debut winner going seven panels here Feb. -

The SOMMET DE L'élevage More Than Ever! the 7, 8 and 9 October
PRESS RELEASE | June 2020 Clermont-Ferrand, France The SOMMET DE L’ÉLEVAGE more than ever! The 7, 8 and 9 October 2020 Today, the SOMMET DE L’ELEVAGE, Europe’s foremost livestock show, can confirm that this year’s edition will be open for business, the 7, 8 & 9 October in Clermont-Ferrand, France. The show’s organisers, while continuing to monitor the situation, note the recent progress in the fight to overcome the COVID-19 epidemic in France. As a result of which, the SOMMET DE L’ELEVAGE will be open for business on the scheduled dates of 7, 8 & 9 October 2020. “We are currently working hard and hand in hand with the local and national authorities as to the measures that we need to put in place to guarantee the health and safety of all the participants”, as Fabrice Berthon, the show’s Director, highlighted. “Though these conditions are not final at this point A PACKED PROGRAMME in time, we will, as soon as possible, be informing all exhibitors and all our visitors.” Beef cattle - the Aubrac breed will hold its national championships at the SOMMET in 2020 with over 400 of the Today, as almost 90% of trade exhibitors have confirmed their presence breed’s best animals in judging, plus an auction sale on Friday 9 October of some of the Aubrac’s best breeding stock. 15 beef at the show, the organisers, despite the difficult context, are already cattle breeds, of which 5 are foreign breeds, will be in judging or confident that this 2020 edition will be good. -

Cob Or Chimera?
Cob or Chimera? UK Expat Sarah Redstone Lee explains why she believes the Cob Normand to be the most versatile cob of them all nce we had decided to take the The origins of the Normandy Cob can be To this day, the Chateau of Versailles is still plunge, sell up, pack up and traced back to Great Britain, the first ‘Norman’ surrounded by carriage driving Normandy make the move to Normandy; horse, to arrive on our shores was during the Cobs in homage to his passion for the breed. code named Operation transportation of a herd of around 2- 3,000, in The National Stud and home of the Overseas ( as but a small preparations for the battle of Hastings of 1066. Normandy Cob was founded by Napoléon in homage to operation Overlord in Normandy, Much of the success of the invasion depended 1806. By 1840, some of the cobs were crossed Owhich formed part of the D-Day landings), it on the horses that were bred for the endurance with King Henry the VIII’s favourite breed, was not long before my husband and I were required in order to carry the weight of the now extinct Norfolk trotter. This further enjoying la belle vie Francaise. We have been armoured suits, weapons and chain mail, yet the enhanced the look of elegance combined here for almost 10 years now, and, aside from superior among them also possessed great speed. with speed. The standard was set and the benefitting from the usual attractions that The Normans, are even to this day well Normandy Cob was then divided in to two France has to offer, that needs no form of known for their breeding of the Percheron groups; heavier horses for work, and lighter introduction, I have been very privileged to and the French Trotter. -

REGLEMENT DU STUD-BOOK DU CHEVAL AUXOIS Article 1 Le
Règlement approuvé le 8 novembre 2011 Pour le ministre et par délégation : Le sous-directeur du développement rural et du cheval Patrick FALCONE REGLEMENT DU STUD-BOOK DU CHEVAL AUXOIS Article 1 Le présent règlement fixe les conditions d’inscription au stud-book du cheval Auxois ainsi que les normes de qualification applicables aux reproducteurs. Il est établi par la commission du stud-book et approuvé par le Ministre chargé de l’agriculture après avis de la commission du livre généalogique. L’établissement public « Institut français du cheval et de l’équitation » (IFCE) est chargé de son application. Article 2 Le stud-book du cheval Auxois comprend : 1) Un répertoire des étalons approuvés pour produire dans la race. 2) Un répertoire des juments destinées à produire dans la race. 3) Un répertoire des poulains inscrits à la naissance au stud-book de la race. 4) Un répertoire des animaux inscrits à titre initial. 5) Une liste des naisseurs de chevaux Auxois. Lors de l’édition périodique du stud-book, n’apparaissent que les reproducteurs ayant eu un produit inscriptible au stud book dans la période de référence. Article 3 Sont seuls admis à porter l'appellation cheval Auxois les animaux inscrits au stud-book du cheval Auxois. Article 4 Les inscriptions au stud-book du cheval Auxois se font au titre de l'ascendance ou à titre initial. Article 5 Inscription au titre de l'ascendance 1) Sont inscrits automatiquement à la naissance au titre de l'ascendance les produits nés en France : a) Issus d’une saillie régulièrement déclarée d’un étalon approuvé. -

Electronic Supplementary Material - Appendices
1 Electronic Supplementary Material - Appendices 2 Appendix 1. Full breed list, listed alphabetically. Breeds searched (* denotes those identified with inherited disorders) # Breed # Breed # Breed # Breed 1 Ab Abyssinian 31 BF Black Forest 61 Dul Dülmen Pony 91 HP Highland Pony* 2 Ak Akhal Teke 32 Boe Boer 62 DD Dutch Draft 92 Hok Hokkaido 3 Al Albanian 33 Bre Breton* 63 DW Dutch Warmblood 93 Hol Holsteiner* 4 Alt Altai 34 Buc Buckskin 64 EB East Bulgarian 94 Huc Hucul 5 ACD American Cream Draft 35 Bud Budyonny 65 Egy Egyptian 95 HW Hungarian Warmblood 6 ACW American Creme and White 36 By Byelorussian Harness 66 EP Eriskay Pony 96 Ice Icelandic* 7 AWP American Walking Pony 37 Cam Camargue* 67 EN Estonian Native 97 Io Iomud 8 And Andalusian* 38 Camp Campolina 68 ExP Exmoor Pony 98 ID Irish Draught 9 Anv Andravida 39 Can Canadian 69 Fae Faeroes Pony 99 Jin Jinzhou 10 A-K Anglo-Kabarda 40 Car Carthusian 70 Fa Falabella* 100 Jut Jutland 11 Ap Appaloosa* 41 Cas Caspian 71 FP Fell Pony* 101 Kab Kabarda 12 Arp Araappaloosa 42 Cay Cayuse 72 Fin Finnhorse* 102 Kar Karabair 13 A Arabian / Arab* 43 Ch Cheju 73 Fl Fleuve 103 Kara Karabakh 14 Ard Ardennes 44 CC Chilean Corralero 74 Fo Fouta 104 Kaz Kazakh 15 AC Argentine Criollo 45 CP Chincoteague Pony 75 Fr Frederiksborg 105 KPB Kerry Bog Pony 16 Ast Asturian 46 CB Cleveland Bay 76 Fb Freiberger* 106 KM Kiger Mustang 17 AB Australian Brumby 47 Cly Clydesdale* 77 FS French Saddlebred 107 KP Kirdi Pony 18 ASH Australian Stock Horse 48 CN Cob Normand* 78 FT French Trotter 108 KF Kisber Felver 19 Az Azteca -

Download File
ENDANGERED BREEDS 1 ENDANGERED BREEDS SISUKORDcontents SISUKORD Estonian Native Cattle .............................................................................................................................................. 4 General ............................................................................................................................................................... 4 History................................................................................................................................................................. 4 Estonian Native Cattle today ....................................................................................................................... 8 Estonian Native Horse .............................................................................................................................................. 9 General ............................................................................................................................................................... 9 A hard working and hardy horse ................................................................................................................ 9 Famous already in ancient times ..............................................................................................................10 Great labour force .........................................................................................................................................12 Estonian Native Horse today .....................................................................................................................14 -

Oesc-Annuaire-Ecus-2016.Pdf
RÉFÉrences - Réseau Écono mique d e la Fil ière Équine ANNUAIRE écusTableau économique, statistique et graphique du cheval en France 2016 Données 2015/2016 Élevage Commerce Utilisations Statistiques socioéconomiques Institut Français du Cheval et de l’Équitation Département diffusion - Librairie Les écuries du Bois - 61310 Le Pin au Haras Tél. : 02 33 12 12 27 • Fax : 02 33 39 37 54 e-Mail : [email protected] • Web : www.ifce.fr Prix public : 10 € TTC © Institut français du cheval et de l'équitation, Novembre 2016. Toute reproduction des statistiques et informations diffusées dans cet annuaire ECUS est autorisé sous réserve de l’indication de la source des données et de la mention du présent document. ISSN : 0760-0488 RÉFÉrences - Réseau Économique de la Filière Équine Présentation Equidés, emplois, entreprises, chiffres d'affaires, répartitions par marché ou territoire, retrouvez l'essentiel sur l'industrie hippique dans ce nouvel annuaire ECUS 2016 ! Depuis sa première parution en 1980, ce document rassemble les chiffres connus de la filière équine et les tendances sur les 10 dernières années. Cet annuaire illustre la richesse des informations existantes en matière de connaissance économique de la filière. C'est une source majeure pour aider à la prise de décision, dans un contexte économique et sociétal évolutif. Cette édition 2016 présente de nouvelles pages sur les résultats économiques des entreprises équines, fruit du partenariat signé en 2015 entre l'Ifce et le réseau Equicer. Elle comporte également de nouvelles données sur l'emploi, les métiers et les formations, extraites du récent rapport national publié par Equiressources. Les pages habituelles sur l'élevage ont été remaniées, pour s'adapter et valoriser au mieux les données disponibles dans la base SIRE.